





(C) 2011 John G. E. Lewis. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
The subgenus Cryptops may be divided into those species with, and those without, an anterior transverse suture on tergite 1. The latter group may be further separated intospecies with the ultimate leg femur without a saw tooth or teeth (here termed the hortensis group) and those with one or more (here termed the doriae group).The species of the Old World of the hortensis group are the subject of this paper. Where possible, the non-European type material has been examined and data for European species added from the literature. Cryptops omissus Ribaut, 1915 (Kenya), Cryptops mirus Chamberlin, 1920 (Tahiti) and Cryptops arapuni Archey, 1922 (New Zealand), lacking ultimate legs cannot be assigned to either the hortensis or the doriae group. Cryptops inermipes Pocock, 1888, which lacks saw teeth on the ultimate legs is considered here. Cryptops nanus Attems, 1938 (Hawaii), Cryptops navis Chamberlin, 1930 (Singapore), Cryptops philammus Attems, 1928 (Southern Africa), Cryptops sinesicus Chamberlin, 1940 (China), Cryptops stupendus Attems, 1928 (South Africa) and Cryptops tahitianus Chamberlin, 1920 (Tahiti) appear from their descriptions to belong to the hortensis group but have a prefemoral saw tooth or teeth and thus belong to the doriae group.
The following species are regarded as valid: Cryptops covertus Chamberlin, 1951, Cryptops daszaki Lewis, 2002, Cryptops decoratus Lawrence, 1960, Cryptops hortensis (Donovan, 1810), Cryptops inermipes Pocock, 1888, Cryptops lobatus Verhoeff, 1931, Cryptops mauritianus Verhoeff, 1937, Cryptops melanotypus Chamberlin, 1941, Cryptops nigropictus Takakuwa, 1936, Cryptops parisi Brolemann, 1920, Cryptops songi Song et al., 2010 and Cryptops stabilis Chamberlin, 1944. Examination of further material is required to establish the true status of Cryptops brignolii Matic, 1977, currently regarded as valid. Cryptops hortensis atlantis Pocock, 1891 is returned to full specific status as Cryptops atlantis.
Cryptops basilewskyi Matic & Darabantu, 1977, Cryptops kalobensis Goffinet, 1971 and Cryptops niloticus Lewis, 1967 are junior subjective synonyms of Cryptops nigropictus. Cryptops penicillatus Lawrence, 1960 is a junior subjective synonym of Cryptops decoratus and Cryptops decoratus sensu Lewis (from Mauritius and Seychelles) is a junior subjective synonym of Cryptops melanotypus. Cryptops parisi cottarellii Matic, 1977, and Cryptops parisi rhenanus Verhoeff, 1931 are junior subjective synonyms of Cryptops parisi. Cryptops campestris Attems, 1953, Cryptops compositus Chamberlin, 1952, Cryptops medius Verhoeff, 1901, Cryptops orthus Chamberlin, 1951, Cryptops parisi sebini Verhoeff, 1934 and Cryptops philammus gabonensis Demange, 1968 are regarded as nomina dubia.
A key is provided to the fourteen species considered valid. However, the status of four of them, namely Cryptops nigropictus, Cryptops decoratus , Cryptops melanotypus and Cryptops songi, which are very similar, and the species that have been here synonymised with them, is uncertain. More material and more refined techniques are required to ascertain their exact status. Whatever that is, however, there can be little doubt that we are dealing with some closely related and very widely distributed populations. This suggests that Cryptops species are easily dispersed over large distances.
Chilopoda, Cryptops hortensis, group, Old World, review, key
The subgenus Cryptops (Cryptops) Leach 1815 may be conveniently divided into those species with, and those without, an anterior transverse suture on tergite 1. The latter group may be further separated intospecies with the ultimate leg femur without a saw tooth or teeth (here termed the hortensis group) and those with one or more (here termed the doriae group). This subdivision may well not be a natural one but is convenient for the present, bearing in mind the parlous state of Cryptops taxonomy.
The hortensis group of Cryptops is the subject of the present paper. It is characterised by a cephalic plate without complete paramedian sutures and generally overlapped by tergite 1, tergite 1 without sutures, and no saw teeth on the femur of the ultimate leg. Cryptops inermipes Pocock, 1888, characterised by the complete lack of saw teeth on the ultimate legs, is also considered here.
Cryptops (C.) zelandicus Chamberlin, 1920, which would appear from the description to belong to the hortensis group, was synonymised under Cryptops spinipes Pocock, 1891 by
The following species: Cryptops omissus Ribaut, 1915 (Kenya), Cryptops mirus Chamberlin, 1920 (Tahiti) and Cryptops arapuni Archey, 1922 (New Zealand), have no anterior transverse suture on tergite 1 but cannot be assigned to either the hortensis or the doriae groups as they lack ultimate legs.
Some species that would appear from their descriptions to belong to the hortensis group in fact have a prefemoral saw tooth or teeth and thus belong to the doriae group. They are: Cryptops nanus Attems, 1938 (Hawaii) (
Specimens were studied by light microscopy using an eyepiece graticule to make drawings onto squared paper calibrated for different magnifications using a stage micrometer. They were examined by reflected light in the preservative (70 or 80% ethanol) and sometimes blotted off briefly to observe surface features such as sulci. They were cleared in 2-phenoxyethanol which was also used as mounting medium for temporary mounts.
The Old World in the title of this paper is taken to mean
the Palaearctic, Afrotropic, Afrotemperate and Oriental regions as
defined by
Conventions and abbreviations used are: [d] indicates that the antenna is damaged, i.e. that it has some articles missing, [r] indicates that the antenna is regenerated.
Uncommon numbers are given in parentheses.
Acronyms
ANSP Academy of Natural Sciences, Philadelphia, Pennsylvania, USA.
BMNH Natural History Museum, London, U.K.
MNHN Muséum national d’Histoire naturelle, Paris, France.
NMNH National Museum of Natural History, Smithsonian Institution, Washington DC, USA.
NMW Naturhistorisches Museum, Vienna, Austria.
MRAC Musée Royal de l’Afrique Centrale, Tervuren, Belgium.
The following taxa are considered here:
Cryptops basilewskyi Matic & Darabantu, 1977
Cryptops brignolii Matic, 1977
Cryptops campestris Attems, 1952
Cryptops compositus Chamberlin, 1952
Cryptops covertus Chamberlin, 1951
Cryptops daszaki Lewis, 2002
Cryptops decoratus Lawrence, 1960
Cryptops hortensis hortensis (Donovan, 1810)
Cryptops hortensis atlantis Pocock, 1891
Cryptops inermipes Pocock, 1888
Cryptops kalobensis Goffinet, 1971
Cryptops lobatus Verhoeff, 1931
Cryptops mauritianus Verhoeff, 1937
Cryptops medius Verhoeff, 1901
Cryptops melanotypus Chamberlin, 1941
Cryptops nigropictus Takakuwa, 1936
Cryptops niloticus Lewis, 1967
Cryptops orthus Chamberlin, 1951
Cryptops parisi parisi Brolemann, 1920
Cryptops parisi cottarellii Matic, 1977
Cryptops parisi rhenanus Verhoeff, 1931
Cryptops parisi sebini Verhoeff, 1934
Cryptops penicillatus Lawrence, 1960
Cryptops philammus gabonensis Demange, 1968
Cryptops songi Song et al., 2010
Cryptops stabilis Chamberlin, 1944
Review of specieshttp://species-id.net/wiki/Cryptops_basilewskyi
Figs 1–6BMNH. 891/V P. and M. Ashmole, St Helena, September to December 2003 (18 mm); 6269/V Prosperous Bay 21.ii.06, P. Ashmole (17 mm).
P. Ashmole’s collection. 1756/V (14 mm); 67/V (16 mm); 452/V (18 mm); 1615/V (18 mm); P and M. Ashmole, St Helena, September to December 2003.
The type material of Cryptops basilewskyi was described from Prosperous Bay, St Helena by
Maximum length 18 mm (14 mm). Colour: cephalic plate light orange, trunk light yellow with dark grey or black pigment on either side of heart and a narrow lateral strip on tergites 3–20, very little on 2 and 21. Pigmented areas also ventrally on 2–6 (Matic & Darabantu, fig.142 a, b, c).
Two basal articles of antenna with long and medium setae changing through 3 and 4 to dense short setae with basal whorl of medium setae on 5. Cephalic plate almost circular the posterior margin covered by tergite 1 (Fig. 1) (not covered by T1). Two very short anterior oblique sutures and short posterior paramedian sutures seen in one specimen only (anterior and posterior sutures present). Clypeus with 2 post-antennal setae, 3 in mid region except for 1615/V where there are 4. Prelabral setae 6. Specimen 6269/V with 2 post-antennal and 5 intermediate setae (Fig. 2).
Forcipular coxosternite very slightly curved on each side with 2 or 3 long to moderate setae (3+3), the innermost slightly further from the edge, and several small setae just behind anterior margin on each side (Fig. 3). Poison gland calyx subspherical situated in anterior region of trochanteroprefemur.
Tergite 1 without sulci, T2 with weak paramedian sulci occupying posterior third to half of tergite, complete 3 or 4 to 20. Lateral crescentic sulci on 3 to 19 or 20. Paramedian sutures could not be seen clearly in cleared specimens. Sternites with median longitudinal sulcus from 2 to 19. Weak transverse sulci 2 to 19 or 20 (only a slightly curved transverse sulcus). Sternite 21 wider than long, posterior margin curved (Fig. 4).
Pore field occupying anterior 54 to 57% of coxopleuron, with 12 or 13 pores (5 to 6 in each field). Three to 6 minute setae in pore field in the 2 largest specimens (no setae), 5 to 6 posterior to the pore field and 5 to 8 on the posterior edge (Figs 4, 5).
Ultimate legs with fine setae except ventrally and posteriorly on the prefemur and ventrally on the femur where they are thicker. Without tubercles or spinous processes. Tibial saw teeth 6 or 8, tarsal 3 (6+4). Tarsal claw without pretarsal accessory spurs.
Legs 1 to 19 with fine setae and tarsus very weakly divided or not (uniarticulate in holotype, biarticulate in paratypes). With a single accessory spur about half length of claw (Fig. 6).
Cryptops basilewskyi has only been recorded from St Helena. It is very similar to Cryptops nigropictus Takakuwa, 1936 from Taiwan and the Ryuku Islands apart from the apparently the faint central longitudinal sulcus on tergites 1 and 2 in the latter. This I regard as trivial character and consider Cryptops basilewskyi to be a junior subjective synonym of Cryptops nigropictus (see p. 35).
http://species-id.net/wiki/Cryptops_basilewskyi
Fig. 7(
Turkey.
Cryptops brignolii is, according to
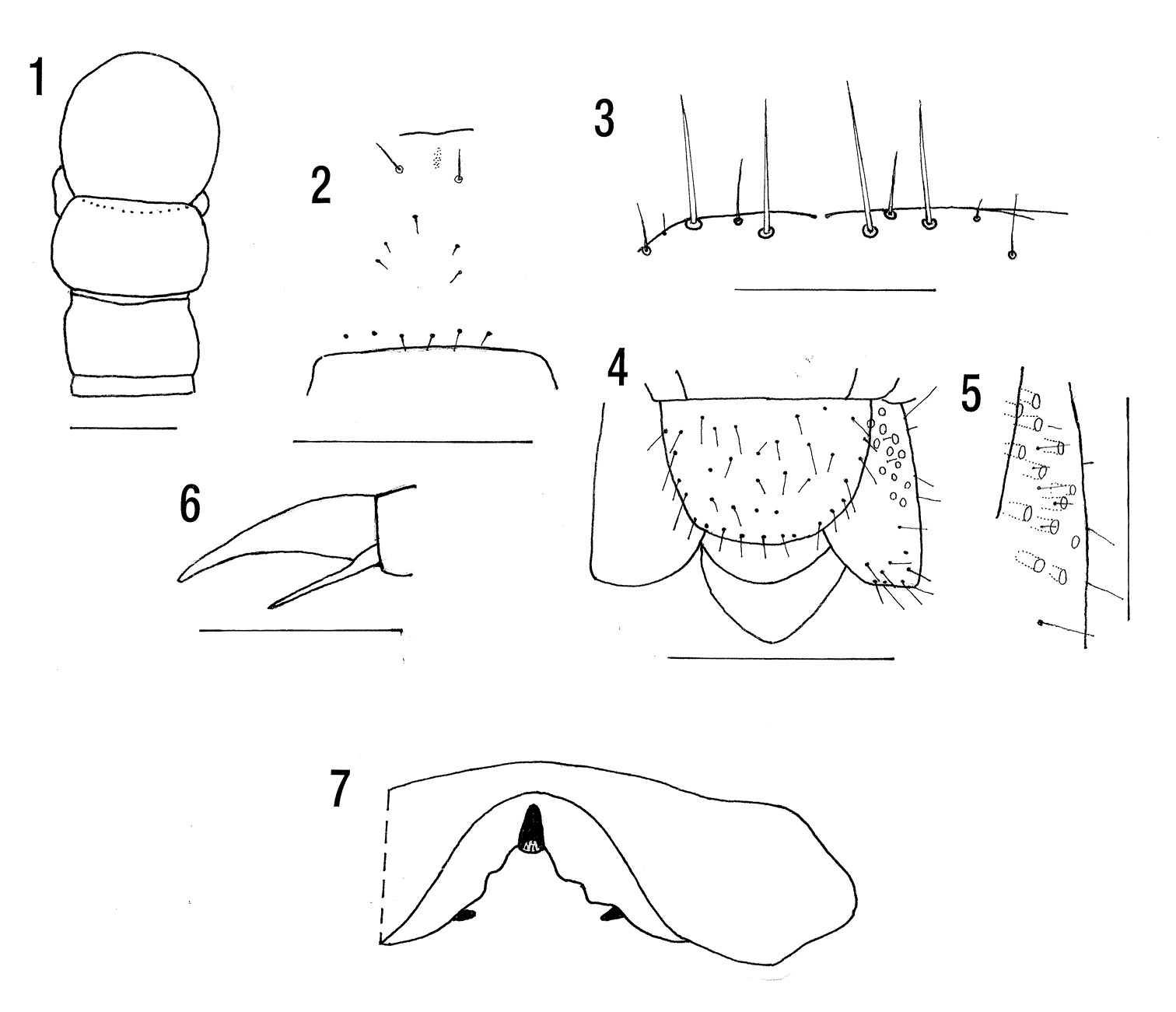
Cryptops basilewskyi. 1 Cephalic plate and tergites 1 and 2 (specimen 1615/V) 2 Clypeus (specimen 6269/V) 3 Anterior margin of forcipular coxosternite (specimen 6269/V) 4 Terminal segments, ventral (specimen 1615/V) 5 Detail of coxopleuron 6 Pretarsus leg 2 (specimen 425/V). Cryptops brignolii. 7 Labrum after
http://species-id.net/wiki/Cryptops_campestris
Figs 8–16NMW. Parts of four syntypes on two slides Inv. Nr. 2950 Cryptops hortensis campestris. Rwanda, Rugege Wald [Forét de Rugege, 2, 000 m]. Slide 1. Cephalic plate, forcipules and anterior segments of three specimens. Slide 2. Cephalic plate, maxillae, forcipules and ultimate legs. There appear to be no complete specimens of this “subspecies” although Attems recorded it from five localities.
(Attems’ (1952) data in parentheses). (Length 12 mm. Colour ochre yellow). Antennal articles short, the basal 3 with some long setae, 3 with some shorter setae, 4 and subsequent with shorter setae and median whorl of long setae. (Head plate and tergite 1 without sutures, the head overlapped by tergite 1). Clypeus with 2+1+2+2 long and 2+2 minute setae (Fig. 8) or 2+1+2 long and 5 minute to small setae and 6 prelabral setae. Anterior wall of coxosternite almost straight, with 1 or 2 long fine setae and 2 or 3 small setae on each side (Figs 9, 10) (with 2+2 large and 2+2 or 2+3 small setae somewhat behind the wall). Poison gland calyx subspherical, situated in anterior region of forcipular trochanteroprefemur (Fig. 11).
(Paramedian sulci from tergite 6, lateral crescentic sulci from 4–19. Tergite 21 without a depression. Sternites with cross sulci).
(Coxopleural pore field reaches hind wall [of?], with some setae). Ultimate legs (Fig. 12) with fine setae on tibia and tarsi and dorsally on prefemur and femur, thicker ventrolaterally, ventrally and ventromedially on prefemur (Fig. 13) and less so ventrolaterally and ventromedially on femur (Fig. 13). Prefemur without median glabrous strip. Tibia with 5, tarsus 1 with 3 saw teeth (Figs 14, 15) (6+3). Legs with long fine setae, pretarsal accessory spurs small (Fig. 16).
Rwanda; Democratic Republic of Congo.
The species has been inadequately described
there being no data on the shape of sternite 21 or the arrangement of
coxopleural pores, but, as
http://species-id.net/wiki/Cryptops_compositus
Turkey.
Chamberlin gave only sketchy data viz. length:
20 mm, cephalic plate and tergite 1 without sutures, tergites with
paramedian sutures. Pore field with up to 15 pores and extending to
posterior margin of coxopleuron. Ultimate legs without spine-like setae
or teeth; with 10 tibial and 4 tarsal saw teeth.
http://species-id.net/wiki/Cryptops_covertus
Figs 17–23NMNH. Four “syntypes” Cryptops covertus Ch. ANG 817.2. [Dundo, Forest of Luachimo, Angola]. Specimen 1: 16 mm, is the best preserved specimen and corresponds most closely with Chamberlin’s description. It is here designated the lectotype, the identity of the other specimens being in doubt. They are: specimen 2: 14 mm (ultimate legs missing), specimen 3 (posterior half only), specimen 4: 9 mm. Specimens 2 and 3 are clearly other species. Specimen 4 was not examined in detail. Chamberlin examined 9 specimens in all.
Length: 16 mm. Without dark subcuticular pigment. Cephalic plate without sutures. Anterior margin of forcipular coxosternite bilobed with 5 or 6 stout marginal setae on each side. Poison gland calyx subspherical. Coxopleural pore field, with about 25 pores and several small setae, occupying no more than anterior 74% of coxopleuron. Ultimate legs with 10 tibial and 3 tarsal saw teeth. Legs 1–20 with short pretarsal accessory spurs.
(
Length 16 mm. Light orange, without subcuticular pigment but with dark stellate bodies beneath the cuticle, presumably fungal (colour light brown, the cephalic plate and last legs a brighter yellow).
Each antenna of 7 long articles probably as the result of elongation after articles lost as suggested by
Tergites with paramedian sulci from 3, lateral crescentic sulci from 4 but with traces on tergites 2 and 3. Sternite cruciform sulci not observed (sulci forming a cross in the middle).
Coxopleuron with about 25 pores of varying size (Fig. 19), the pore field, with several small spine like setae, occupying the anterior 74% of coxopleuron. With 6 setae on the posterior margin and a single seta just anterior to them. Prefemur of ultimate leg (Fig. 20) with fine setae dorsally and dorsolaterally, with spine-like setae (Fig. 21) ventrolaterally, ventrally and medially except for a longitudinal glabrous area medially. Femur with fine setae dorsally, spine-like setae (Fig. 21) and a few fine setae ventrally and spine like setae ventromedially. Tibia and tarsus 1 and 2 with fine setae. Right tibia with 10 saw teeth, comprising 8 and a second row of 2, tarsus with 3 saw teeth (Fig. 22) (Tibia with 4 stout teeth in series below [meaning?], the first tarsal joint with 2. Tibia with a low obtuse tooth on each side at distal end above). Legs 1–19 with undivided tarsi and fine setae. Pretarsi of legs 1–20 with short accessory spurs (Fig. 23).
Angola.
Chamberlin’s description of the arrangement of saw teeth on the ultimate legs differs markedly from that of the lectotype which otherwise compares well with his brief description. Further material is required to resolve this difference.
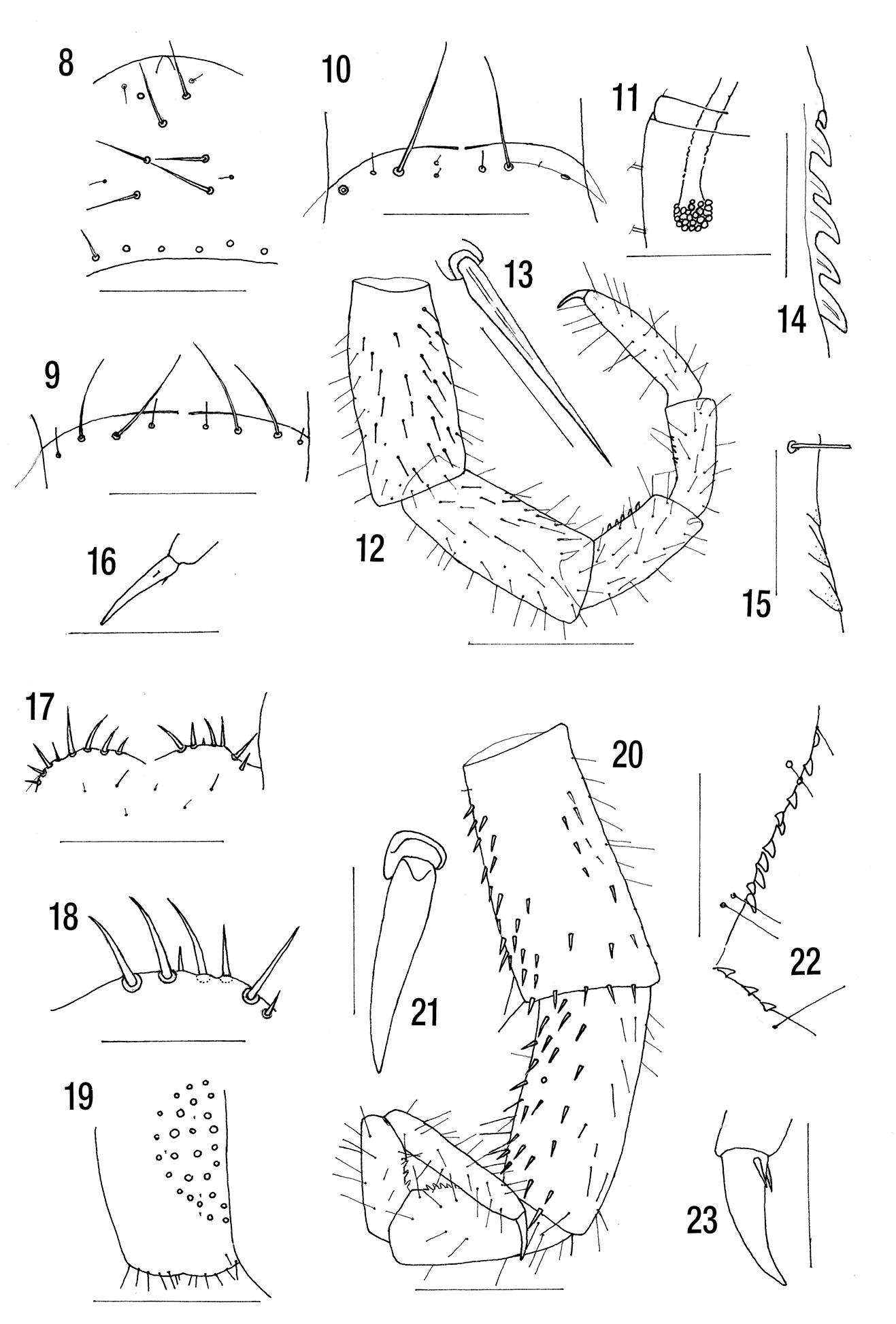
Cryptops campestris syntypes. 8 Clypeus 9 Anterior margin of forcipular coxosternite (slide 1) 10 Anterior margin of forcipular coxosternite (slide 2) 11 Poison gland calyx (slide 2) 12 Ultimate leg (slide 2) 13 Ventromedial spine from ultimate prefemur 14 Tibial saw teeth ultimate leg 15 Tarsal saw teeth ultimate leg 16 Pretarsus leg 3 (slide 2). Cryptops covertus lectotype17 Anterior margin of forcipular coxosternite 18 Detail of left half of forcipular coxosternite 19 Ultimate leg coxopleuron 20 Ultimate leg median 21 Spine-like seta from ultimate prefemur 22 Tibial and tarsal saw teeth ultimate leg 23 Pretarsus of leg 16. Scale lines = 0.1 mm except Figs 12, 19, 20 = 0.5 mm, Figs 17, 22 = 0.25 mm and Figs 13, 21 = 0.05 mm.
http://species-id.net/wiki/Cryptops_daszaki
Figs 24–27Length: 4–7 mm. Without dark subcutaneous pigment. Cephalic plate without sutures. Tergite paramedian sutures complete from 3. Anterior margin of forcipular coxosternite slightly concave or straight with 2 or 3 fine setae on or almost on anterior margin. (Fig. 24). Poison gland calyx with very few gland cells, situated in anterior part of forcipular trochanteroprefemur (Fig. 25). Sternite almost semicircular (Fig. 26). Pore field occupying 50% of coxopleuron and with 5 or 6 pores. Ultimate leg with 3 tibial and 2 tarsal saw teeth. Pretarsi of legs 1–20 with a single long accessory spur. (Fig. 27).
Île aux Aigrettes, Mauritius (Indian Ocean).
Some specimens, at least, of this very small species were mature and
http://species-id.net/wiki/Cryptops_decoratus
Figs 28–30MNHN. Holotype [Madagascar Sambirano]. Mt. Tsaratanana [Andilabe] 2200 m. II 51 RP [R. Paulian, févr, 1951]. (Forcipules are missing).
(Based on Cryptops decoratus sensu
(Additional data from
Length 14 mm. Colour after preservation for 48 years uniform greyish orange (yellow with green black pigment forming a central double band and along lateral margins from tergite 2).
Cephalic plate without sutures. Tergite 1 without
sutures and overlying posterior margin of cephalic plate. (Anterior
margin of coxosternite almost straight with 1+2 long submarginal and 1+2
shorter setae on each side. Poison gland calyx spherical, situated in
anterior part of forcipular trochanteroprefemur [these data from
Tergites with median ridge from 3 (5–20).
Paramedian sutures from 5 (“sillons” from 5 or 6–20). Lateral crescentic
sulci from 3–20 (5–19). Tergite 21 with posterior median depression.
Sternites with cruciform sulci, the longitudinal sulcus longer and
wider than the narrow lateral arms (the longitudinal sulcus present on
all segments, the transverse weak and absent on anterior half of
trunk). (Sternite 21 with posterior margin straight or slightly
concave). Sternite 21 was not figured by
Coxopleuron with 8 large pores (7–9) occupying anterior 50% of coxopleuron and with at least 3 minute setae in pore field; 3 or 4 fine setae on posterior margin and up to 5 between this and the pore field (several moderately long setae between the pores) (Fig. 28). Prefemur of ultimate legs with long fine setae dorsally, but thicker setae on lateral, ventral and medial surfaces (Fig. 29) without medial longitudinal glabrous area. Femur with long fine setae, those of medial surface thicker. (End legs with numerous strong setae). Tibia with 4, tarsus 1 with 2 saw teeth (5+2/3). Legs 1–19 with fine setae, tarsi undivided, pretarsal accessory spurs very short (Fig. 30).
Madagascar.
Cryptops penicillatus (from Madagascar Centre and Madagascar Sambirano) is very similar to Cryptops decoratus (from Madagascar Sambirano, Madagascar Nord and Madagascar Est). These are distinguished in Lawrence’s (1960) key as follows:
| – | Colour uniform yellow: transverse sulci of sternites stronger than the longitudinal sulci | Cryptops penicillatus |
| – | Greenish black pattern on dorsal surface and sometimes also on the pleural and sternites; longitudinal sulci of sternites clearly more marked than the transverse sulci | Cryptops decoratus |
His key, however, does not tally with the text which states that Cryptops penicillatus is without transverse sulci on the sternites and Cryptops decoratus the transverse sulci are weak. These do not tally with my observations on the two holotypes which both have a broad longitudinal and a narrow transverse sulcus and thus do not differentiate the species. I regard the two as conspecific Cryptops penicillatus being a Cryptops decoratus lacking dark subcutaneous pigment.
NB.
Figs 31–38
(The latter in parentheses where appropriate).
Length: 10–15 (7.5–14) mm. With dark subcuticular pigment, though very
little in specimens from Serpent Island as compared to those from Round
Island (only 5 of 15 specimens from the Seychelles had black or grey
subcuticular pigmentation the remainder showed no dark colouration).
Cephalic plate and T1 without sutures. Clypeus with 2+1+2+6 (Fig. 34)
or 2+1+2+2+6 setae. Anterior margin of forcipular coxosternite slightly
curved on each side with 2+2 or 2+3 or 3+3 long submarginal setae (Fig. 31) (2+2). Poison gland calyx subspherical, situated in anterior region of forcipular trochanteroprefemur (Fig. 32). Posterior margin of sternite 21 straight or slightly curved (mostly with rounded posterior margin, rarely straight) (Fig. 35).
Pore field occupying anterior 60–70% of coxopleuron, with 7–16 pores
(8–14), 5–8 small setae in the pore field (4–6) and 1–3(4) between it
and the coxopleural margin ((0)1–2) (Fig. 36). Ultimate leg prefemora with broad medial longitudinal glabrous area with 5–7 tibial and 3–4 tarsal saw teeth (Fig. 37).
Pretarsal accessory spurs of legs 1–20, the posterior 40% length of pretarsus the anterior slightly shorter but sometimes only 12% of the pretarsal length (Fig. 33). The accessory spurs short in Seychelles material, the second very short and easily overlooked (Fig. 38).
The specimens from Mauritius and the Seychelles may be differentiated from Madagascan Cryptops decoratus by the possession of the glabrous area on the ultimate prefemur and lower number of setae posterior to the coxopleural pore fields. These differences are relatively minor but consistent. They are clearly very closely related to Cryptops melanotypus Chamberlin, 1921 and are here regarded as belonging to that species although it is known only from a single specimen (see p. 3).
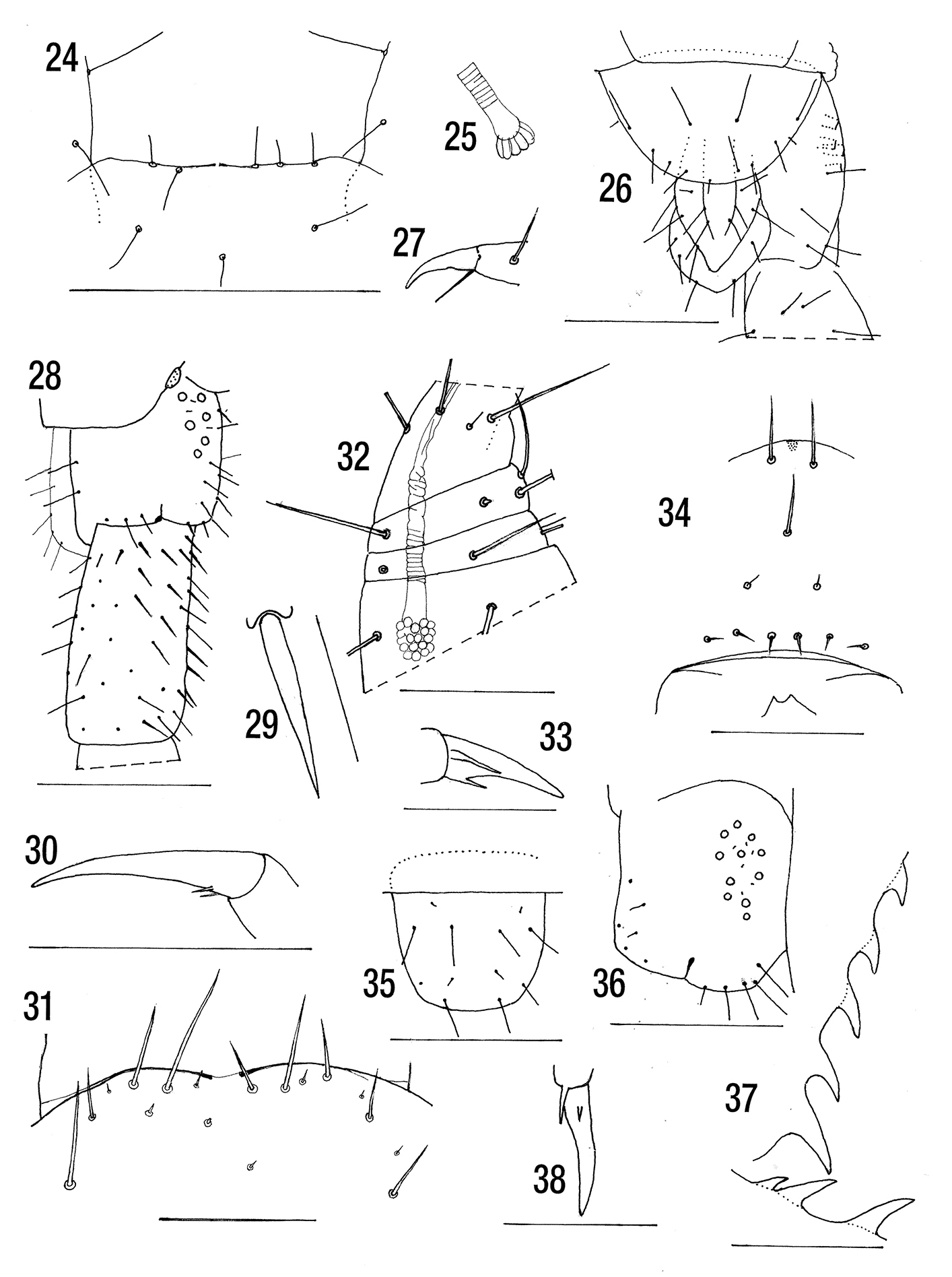
Cryptops daszaki holotype after
http://species-id.net/wiki/Cryptops_hortensis_hortensis
Figs 39–44Length: about 20 mm. Without dark subcutaneous pigment. Cephalic plate with very short anterior longitudinal sutures extending only a short way back from the bases of the antennae, without posterior paramedian sutures. Clypeus with a pair of post-antennal setae, 4–12 somewhat irregular clypeals and 6–12 prelabrals. Labral sidepieces not notched. Anterior margin of forcipular coxosternite barely protuberant, with about four setae on each side on the anterior margin (Fig. 39). Poison gland calyx club-like (Fig. 40), somewhat variable in shape, situated mainly in the forcipular tibia. Posterior margin of sternite 21 slightly concave. Coxopleural pore field with about 30 pores, not extending to posterior margin, usually containing a single seta (Fig. 41), sometimes without. Prefemur of ultimate leg with ventral median longitudinal groove. With 5–9 tibial (Fig. 42) and 2–4 tarsal saw teeth (Fig. 43). Pretarsal accessory spurs of legs 1–20 very small (Fig. 44).
Azores, Madeira, Canary Islands, Morocco, Iberia, France (incl. Corsica), Ireland, United Kingdom, Denmark, Norway, Netherlands, Germany, Poland, Czech Republic, Slovak Republic, Hungary, Italy (incl. Sardinia and Sicily), Switzerland, Austria, Slovenia, Croatia, Bosnia-Herzegovina, Montenegro, FYR Macedonia, Albania, mainland and insular Greece (incl. Crete), Bulgaria, Romania, N-Turkey, Ukraine, European Russia, Georgia, Armenia, Azerbaijan, Turkmenistan, Tajikistan, Uzbekistan; introduced into United States, Hawaii, Canada, St. Helena; Australia (Tasmania), Christmas Island. Synanthropic in Scandinavia (Sweden, Finland).
Cryptops hortensis pauciporus Brolemann, 1908 has no seta in the pore field.
http://species-id.net/wiki/Cryptops_hortensis_atlantis
Figs 45–54BMNH. Holotype. Madeira: Grant, W. R. O. BMNH(E) # 20001 Chilo. 1891.3.8.10. Original label missing.
Length 20.5 mm. Without dark subcutaneous pigment. Cephalic plate with very short longitudinal sutures extending only a short way back from the bases of the antennae. Clypeus with a pair of post-antennal setae, 12 somewhat irregular clypeals and 9 prelabrals. Labral sidepieces not notched. Anterior margin of forcipular coxosternite barely protuberant, with 4 to 6 fine setae on each side on the anterior margin. Poison gland calyx cylindrical, situated mainly in posterior part of the tarsungulum. Posterior margin of sternite 21 slightly concave. Coxopleural pore field not extending to posterior margin, without setae. Prefemur of ultimate leg with ventral median longitudinal groove. Tibia with 15–17 well separated minute saw teeth, tarsus with 5–6 close set teeth. Pretarsal accessory spurs of legs 1–20 very small.
(
Male (contains 3 spermatophores). Length 20.5 mm (21.5 mm). Colour reddish brown (antennae, head, first two and last two somites and anal legs ochraceous; rest of legs testaceous: rest of somites ochraceo-fuscous).
Antennnal articles 15[r]-12[d], article 1 with long and medium setae, 2 with in addition a few short setae, 3 intermediate, 4 with small setae and basal whorl of long and medium setae. Cephalic plate with extremely short weak oblique anterior paramedian sutures at bases of antennae. Tergite 1 overlying posterior margin of cephalic plate.
Clypeus with a pair of post-antennal setae, about 12 clypeals of varying size approximately forming an elongated triangle (Fig. 45). A row of 9 prelabral setae. Labrum not visible.
Anterior wall of forcipular coxosternite slightly convex on each side with 6 + 4 fine marginal setae (Fig. 46). Poison gland calyx situated in tibia and posterior part of tarsungulum, cylindrical and of moderate length (Figs 47, 48).
Tergite sutures not seen, paramedian sutures
difficult to observe in this darkly pigmented specimen but apparently
wanting on tergites 2 and 3, occupying anterior 75% on tergite 4,
complete on 14, almost complete on 19 and 20. Very weak arcuate sutures
on tergites 4 to 8. (The first three wholly without sulci, the fourth
obsoletely sulcate posteriorly and laterally, the rest, except the
last, with four sulci; two internal complete, two external incomplete
and oblique, the oblique sulci almost obsolete on the seventeenth to
twentieth tergites).
Detail of sternites 1 to 20 not observed (medially and longitudinally sulcate, the transverse sulcus scarcely perceptible). Sternite 21 with sides converging posteriorly and hind margin slightly concave (Fig. 49). Coxopleuron with relatively few pores (19–20) occupying anterior 70% of pore field. No setae in pore field and none between pore field and posterior margin which bears 6 or 7 setae.
Ultimate leg (Fig. 50) prefemur with spinous setae on ventral and posterior (median) surface, dorsal and lateral surfaces with a few setae. A glabrous ventromedial groove. Femur with fewer spinous setae ventrally and medially but more fine setae especially distally. A poorly defined glabrous ventromedian strip. Tibia flattened dorsally, and in distal half with a few short fine setae dorsally and laterally and dense fine setae ventrally and medially: with 15 or 17 well-separated minute saw teeth on inferior edge of flattened medial surface (Fig. 51). The attendant setae parallel-sided and pointed (Fig. 52). Tarsus 1 bulbous distally with dense medium to long setae, with 5 or 6 close set saw teeth (Fig. 53), larger than those of the tibia and set on a pronounced hump. Tarsus 2 with a few scattered setae. Pretarsal claw simple, Pretarsal accessory spurs of legs 1–20 small (Fig. 54). The twentieth pair of this male with dense fine setae ventrally on prefemur, femur and tibia.
Madeira and possibly the Azores and the Canary Is.
The differences in the number and type of the saw teeth between Cryptops hortensis atlantis and Cryptops hortensis are major and I consider them more than sufficient to return Cryptops atlantis to full specific status. It is worth noting that
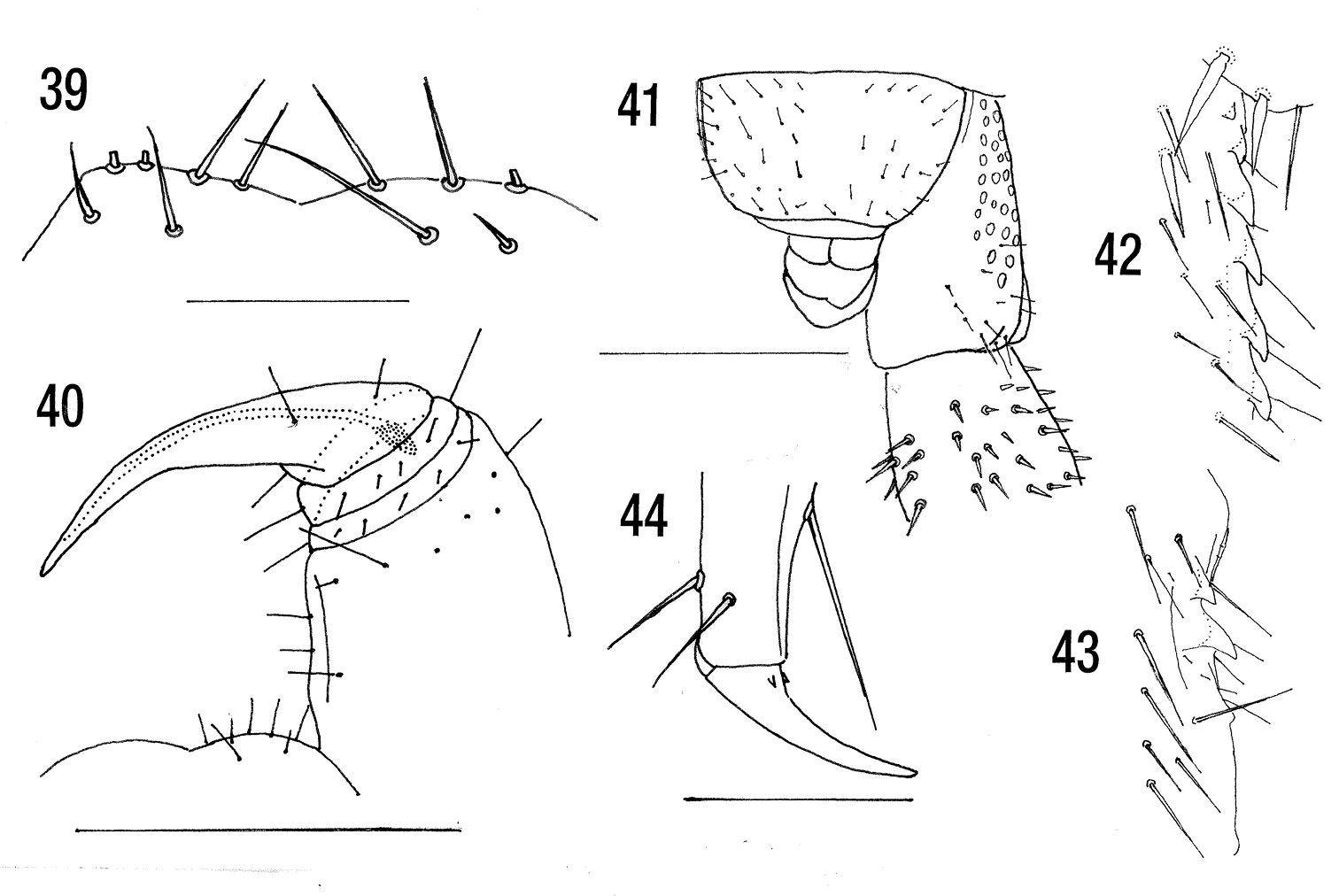
Cryptops hortensis hortensis. 39 Anterior margin of forcipular coxostrernum (St Helena specimen 1) 40 Left forcipule and coxosternite (La Guardia, Spain specimen C17) 41 Terminal segments, left coxopleuron and part of left prefemur of ultimate leg (Bournemouth, U.K.) 42 Tibial saw teeth of ultimate leg (after
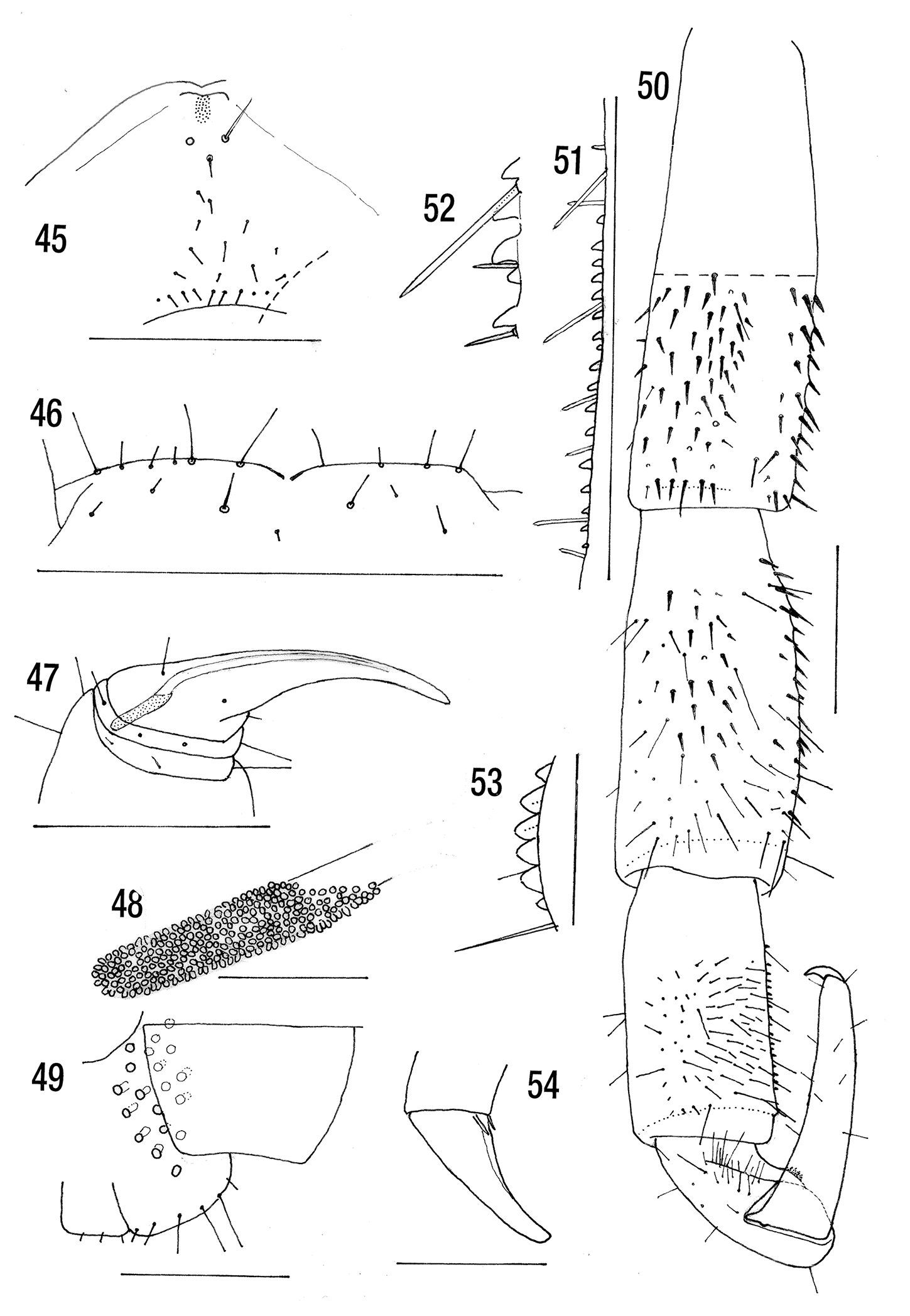
Cryptops hortensis atlantis holotype. 45 Clypeus 46 Anterior margin of forcipular coxosternite 47 Right forcipule 48 Detail of poison gland calyx 49 Sternite 21 and right coxopleuron (sternite setae omitted) 50 Left ultimate leg 51 Ultimate leg tibial saw teeth 52 Detail of saw teeth and setae 53 Ultimate leg tarsal saw teeth 54 Pretarsus of ambulatory leg. Scale lines= 0.5 mm except Figs 48, 51, 53, 54 = 0.1 mm.
http://species-id.net/wiki/Cryptops_inermipes
Figs 55–62BMNH. Holotype labelled Cryptops (Cryptops) inermipes Pocock type Indian Ocean: Xmas Island Lister J J. 88.96 HMS ‘Egeria’ BMNH(E) 200007 Chilo 1888.96.
Length 27 mm.Without dark subcutaneous pigment. Cephalic plate without sutures. Anterior margin of forcipular coxosternite barely protuberant, with three or four fine submarginal setae on each side on the anterior margin. Poison gland calyx ovoid, situated mainly in the anterior part of the trochanteroprefemur. Posterior margin of sternite 21 slightly curved. Coxopleural pore field occupying anterior 75% of coxopleuron, with about 70 pores. Ultimate tibia and tarsus without saw teeth. Pretarsal accessory spurs of legs 1–20 minute.
(
Length 27 mm. Colour: brownish orange, cephalic plate brown. Antennal articles 16 [d]+16 [d], very long. Article 10 twice as long as wide (Fig. 55) with an irregular whorl of long basal setae (Fig. 56).
Cephalic plate without sutures, overlain by first tergite. Forcipular coxosternite slightly convex on each side with 4+3 fine submarginal setae (Fig. 57). Calyx of forcipular poison gland ovoid and situated in the anterior part of the trochanteroprefemur (Figs 58, 59).
Paramedian and crescentic sulci clearly visible from tergite 3 (second [tergite] with two shallow sulci; the rest, with the exception of the last, bearing four sulci). Sternite sulci not seen but the transverse apodeme is clear (each, the first excepted, with a median longitudinal sulcus, and most of them with a median transverse sulcus, the distinctness of this latter sulcus is variable, and it almost disappears in the posterior region of the body).
Sternite 21 with sides converging posteriorly with a slightly convex posterior margin (Fig. 60). Coxopleuron with about 70 pores and occupying approximately anterior 75% of the pore field. With two setae in the pore field, one behind and 6 on the posterior margin.
Ultimate legs now missing (denticles absent upon the under surface of the tibial and proximal tarsal segments of the anal legs). Twentieth pair of legs now missing. Ambulatory legs (Fig. 61) with undivided tarsi (legs, all that remain, remarkably hirsute but none of the segments spiny.). Pretarsal accessory spurs minute (Fig. 62).
Christmas Island (Indian Ocean).
The lack of saw teeth on the ultimate legs would
appear to be an extremely rare mutation. The condition has not been
reported from any other species in the genus. Unfortunately the fact
that the ultimate legs are now missing does not allow confirmation of
the condition. It is possible that
http://species-id.net/wiki/Cryptops_kalobensis
Figs 63–65MRAC. Three paratypes 9, 11 and 12 mm. Det G. Goffinet 1970 Congo: Katanga, Chute Kaloba, rivière Lofoi, base du plateau des Kundulungu [alt 1, 070 m (Freson, R.). Réf. nº CM 1661]. 2.xi.1968. R.G. Mus. Afr. Centr. 13.135
(
I regard Cryptops kalobensis, from the Democratic Republic of Congo, as conspecific with Cryptops nigropictus from Taiwan and the Ryuku Islands of which it is a junior subjective synonym (see p. 35).
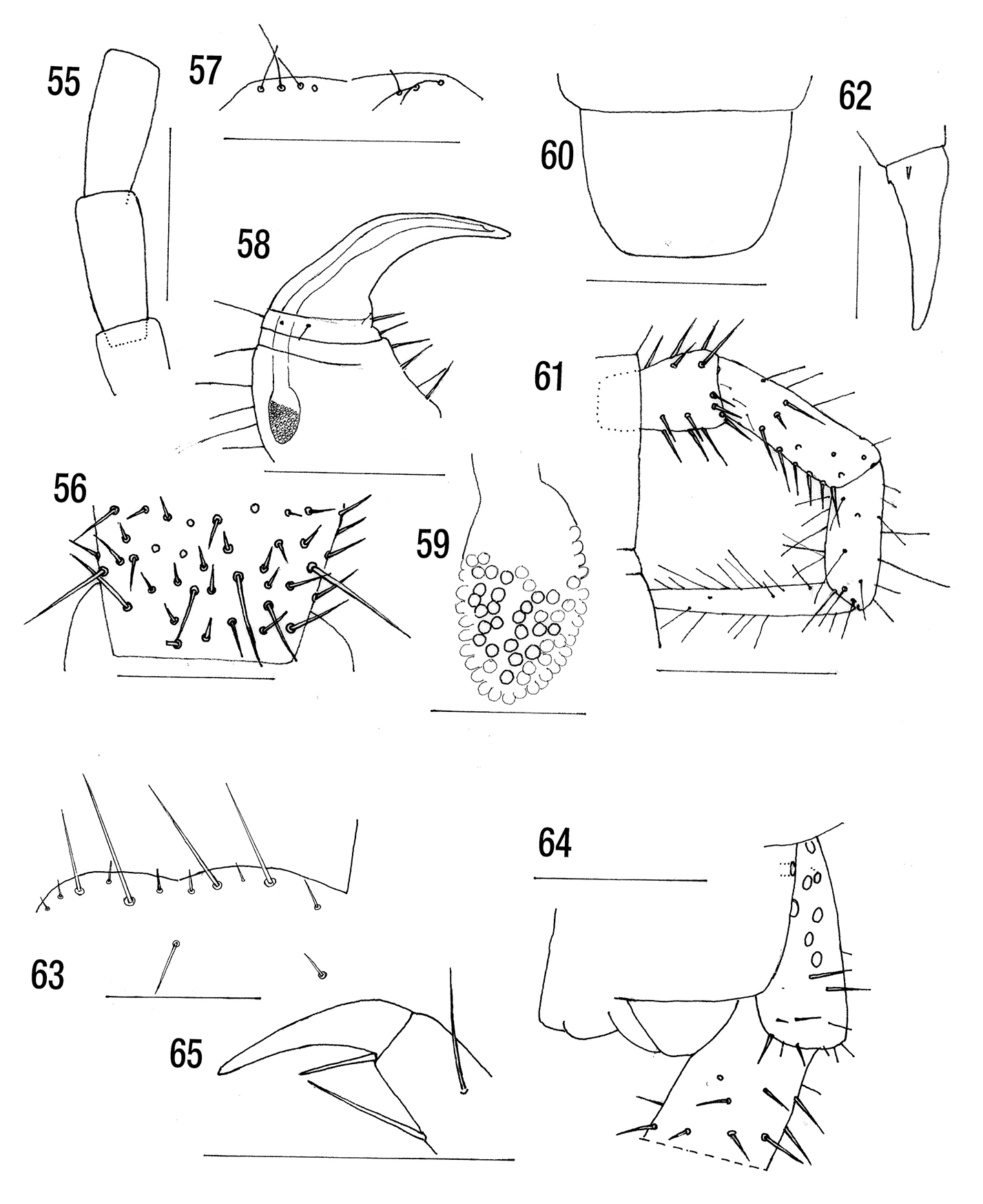
Cryptops inermipes holotype. 55 Antennal articles 10, 11 (setae omitted) 56 Detail of proximal region of antennal article 10 57 Anterior margin of forcipular coxosternite 58 Right forcipule 59 Detail of poison gland calyx 60 Sternite 21 61 Leg 2 62 Pretarsus leg 18. Cryptops kalobensis. 63 Anterior margin of forcipular coxosternite (paratype specimen 2) 64 Coxopleuron and parts of sternite 21 and ultimate leg prefemur (paratype specimen 1) 65 Pretarsus leg 9 (paratype specimen 3). Scale lines = 0.5 mm except Fig. 64 = 0.25 and Figs 56, 59, 61, 63–65 = 0.1 mm.
Cryptops lobatus. 6. Forcipular coxosternite and right forcipule (after
http://species-id.net/wiki/Cryptops_lobatus
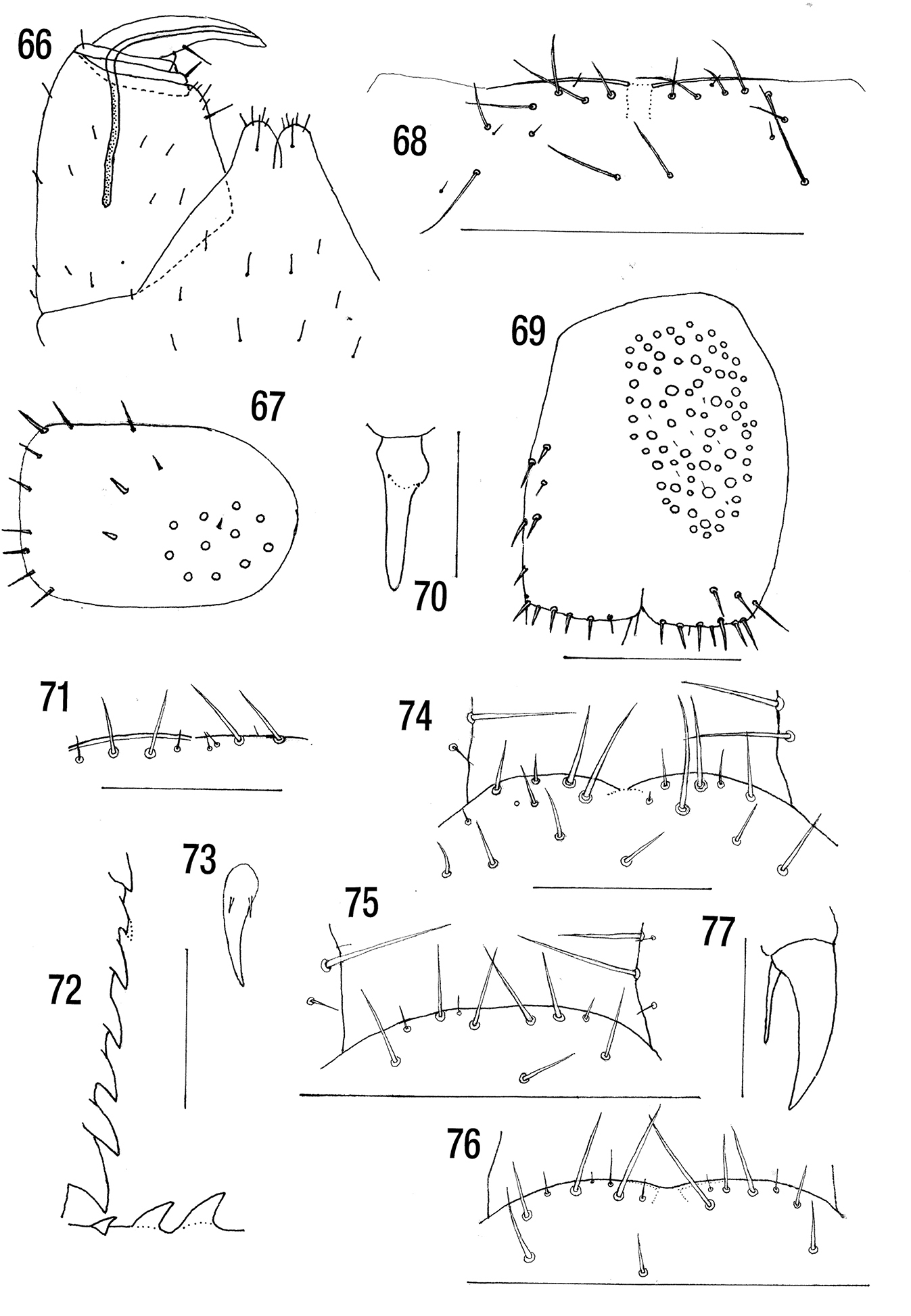
Figs 66, 67 (Based on data in
Southern France, Italy (Liguria).
http://species-id.net/wiki/Cryptops_mauritianus
Figs 68–70Length: 15–32 mm. With dark subcuticular pigment.
Cephalic plate with or without very short
posterior paramedian sutures. Anterior margin of forcipular coxosternite
almost straight with (3) 4 or 5 long submarginal setae on each side (Fig. 68). Not each side with 3+4–4+5 as stated by
Distribution. Mauritius (SW Indian Ocean)
http://species-id.net/wiki/Cryptops_medius
Greece.
http://species-id.net/wiki/Cryptops_melanotypus
Figs 71–73NMNH. Holotype. Cryptops melanotypus C. From Philippines at Honolulu Dec 27, 1937 27–387. [In packing material about Den(robium) superbum from the Philippine Islands].
Length: 9–16 mm. With or without dark subcuticular pigment. Cephalic plate without sutures. Anterior margin of forcipular coxosternite slightly curved and with 2 (3) submarginal setae on each side. Posterior margin of sternite 21 straight or rounded. Coxopleural pore field occupying no more than anterior 60% of coxopleuron, with 6–17 pores, 0–8 setae in coxopleural pore field, 0–3 posterior to it. Ultimate leg prefemur with a longitudinal glabrous area medially. With 4–7 tibial and 3 (4) tarsal saw teeth. Legs 1–20 with two short pretarsal accessory spurs.
(
Length 9 mm. Tergites 3–20 with dark brown (black) pigment on either side of heart and along lateral edges with patches between tergite 21 with faint median posterior patch. Sternites 2–20 with pigment patches.
Antennal articles 10[r]-16. Cephalic plate without sutures, overlappe by tergite 1. Anterior margin of forcipular coxosternite slightly curved on each side, with 2 long and 2 very short submarginal setae on right and 2 long and 3 very short on left. Those on left appear mostly marginal probably due to distortion (Fig. 71).
Tergites with incomplete median ridge from 7–17, lateral crescentic sulci from 3, incomplete paramedian sulci from 6 but these latter very difficult to determine as the cuticle is wrinkled longitudinally as seen in specimens in early stages of ecdysis (first and subsequent tergites bisulcate, the sulci mostly complete). Traces of paramedian sutures seen only on tergite 10 and 15 in the cleared specimen. Sternite cruciform sulci not observed. Sternite 21 with broadly rounded posterior corners, posterior margin straight.
Coxopleuron with 17 pores, 6 beneath the margin of sternite 21, the pore field occupying the anterior 60% of coxopleuron and without setae. Two setae between pore field and posterior margin on which there are 5 setae. Ultimate legs (loose) with long fine setae, a longitudinal glabrous area medially on prefemur. Tibia with 7, tarsus 1 with 3 sharp saw teeth (Fig. 72). Legs 1–19 with long fine setae, tarsi clearly divided from 17, pretarsal accessory spurs short (Fig. 73).
Philippines, Mauritius, Seychelles.
The holotype is small and delicate, nevertheless most characters are visible. The absence of minute setae from the coxopleural pore field may be a juvenile character. It has a relatively high number of coxal pores and ultimate leg saw teeth for such a small specimen. With the current state of our knowledge it is, however, not possible to separate it from the “Cryptops decoratus” populations from Mauritius and the Seychelles, described above, which I regard as conspecific with it and hence junior subjective synonyms.
http://species-id.net/wiki/Cryptops_nigropictus
Length 11–18 mm. Tergites with dark subcuticular pigment. Anterior margin of forcipular coxosternite slightly curved, typically with 2 long and several shorter submarginal setae on each side. Poison gland calyx rounded, situated in anterior part of forcipular trochanteroprefemur. Posterior margin of sternite 21 rounded. Coxopleural pore field with 7–25(31) pores and 0–6 setae in pore field and 3–9 posterior to it. Ultimate leg with 4–8 tibial and 3–4 tarsal saw teeth. Legs 1–20 with a single long pretarsal accessory spur.
(Based on
St. Helena, Democratic Republic of Congo,
Sudan, ?Eritrea, Somalia, Yemen, Mauritius, Rodrigues,
Philippines, China, Taiwan, and Ryuku Islands (Japan). Not French
Polynesia as stated in
The apparent close similarity of Cryptops basilewskyi, Cryptops kalobensis and Cryptops niloticus to Cryptops nigropictus suggests that all four are representatives of a single widely distributed species (see Discussion for a fuller examination of this thesis). Cryptops nigropictus is the senior subjective synonym.
http://species-id.net/wiki/Cryptops_niloticus
Figs 74–77BMNH Holotype BMNH(E) 200011 Chilo 1996.9.62 Sudan 28.ix.1964. Holotype Reg. No. 1966.9.6.2. Khartoum, Sudan 28.ix.1964. Paratype Reg. No. 1966.9.6.3. Khartoum, Sudan 9.ix.1962.
(Sudanese material) Length 16 (male)–20 mm (female). With dark brown subcuticular pigment. Cephalic plate without or with very short posterior paramedian sutures. Anterior margin of forcipular coxosternite typically 2+2(3) and several smaller setae some just submarginal (Fig. 74). Poison gland calyx spherical or subspherical situated in anterior region of forcipular coxosternite. Posterior margin of sternite 21 broadly rounded. Pore field occupying anterior 70% of coxopleuron with 10–25 (31)pores. With 5 setae in pore field, 3 between pore field and posterior margin on which there are 4 or 5 setae (these data from 2 specimens only). Ultimate legs with 4–7 tibial and 3 tarsal saw teeth. Pretarsi of legs 1–20 each with a single long accessory spur 54–66% of length of pretarsus.
The brief description of the specimens from
Eritrea precludes definite confirmation of identity. They were destroyed
in a fire at Ahmadu Bello University, Zaria, Nigeria. A male from the
Yemen (
http://species-id.net/wiki/Cryptops_orthus
(Data from
Angola.
The limited data provided by
http://species-id.net/wiki/Cryptops_parisi_parisi
Figs 78–85 Length 15–25 mm. Without dark subcuticular
pigment. Cephalic plate, with sutures extending only a short way back
from the bases of the antennae and posterior sutures occupying the
posterior third of the cephalic plate (absent in young animals) (Fig. 78). Labral sidepieces notched in most specimens (tridentate labrum) (Fig. 79), sometimes notched on one side only but not notched (unidentate) in some Iberian populations (
Netherlands, Germany, France, Italy (excluding Sardinia and Sicily), Austria, former Czechoslovakia, Slovenia, Croatia, Bosnia-Herzegovina, Montenegro, FYR Macedonia, Albania, Romania, Bulgaria, mainland Greece (including Ionian Is.), N. Turkey, European Russia (Transcarpathia); synanthropic in UK and Ireland, introduced in Scandinavia and Canada (Newfoundland).
http://species-id.net/wiki/Cryptops_parisi_cottarellii
(Data from
This Turkish subspecies is not listed in
http://species-id.net/wiki/Cryptops_parisi_rhenanus
Characterised mainly by the poison gland pore tube reaching almost to the base of the trochanteroprefemur as opposed to about the middle. Probably individual variation and regarded here as a junior subjective synonym of Cryptops parisi parisi. Locality not given but rhenanus means from the banks of the river Rhine.
http://species-id.net/wiki/Cryptops_parisi_sebini
Listed as a valid subspecies by
http://species-id.net/wiki/Cryptops_penicillatus
MNHN. Holotype. [Madagascar Centre: forêt de] Manjakatompo, Ankaratra Mts 2, 000 m, I 1956. (Forcipules are missing).
(Additional data from
Length 16.5 mm. Colour yellow, without dark
subcuticular pigment. Antennal articles 12+11, the basal 2 with long, 3
with, in addition, some short setae. The number of short setae
increase over 4, 5, and 6. An irregular basal whorl of long setae from
4. Cephalic plate without sutures. (Tergite 1 without sutures and
overlying posterior margin of cephalic capsule. Anterior margin of
coxosternite weakly convex with 2+2 long and 3+3 short setae on each
side [these data from
(No paramedian sulci or sutures (“sillons”) before tergite 7, indistinct on the other tergites. Lateral sulci only occupying the anterior third of each tergite, last 3 tergites without sulci). Sternites with deep wide longitudinal sulcus (on all sternites except the first), transverse sulcus very fine (none). Sternite 21 with corners broadly rounded and posterior margin straight (small, rounded).
Coxopleuron with 8 large pores (6–8) occupying anterior 55% of coxopleuron and with at least 6 minute setae in pore field. Six fine setae on posterior margin and 5 between this and the pore field. Ultimate legs with fine setae, the ventrolateral setae of the prefemur thicker, without a medial longitudinal glabrous area. Tibia with 6, tarsus 1 with 4 saw teeth (5+3). Legs 1–19 with fine setae, tarsi undivided, pretarsal accessory spurs very small. (Leg 20 three basal articles with a brush of setae ventrally).
Described from Madagascar, Cryptops penicillatus is very similar to and is hereregarded as a Cryptops decoratus lacking dark subcutaneous pigment of which it is a junior subjective synonym. For a full discussion of this see p. 23.
http://species-id.net/wiki/Cryptops_philammus_gabonensis
(Data from
Gabon.
The data available are insufficient for the exact identity of Cryptops parisi gabonensis to be determined. It is here regarded as a nomen dubium. As noted in the introduction, Cryptops philammus has a saw tooth on the ultimate leg femur and thus belongs to the doriae group of species.
http://species-id.net/wiki/Cryptops_songi
Maximum length 22 mm. Cephalic plate with very short posterior paramedian sutures, anterior margin of coxosternite with 3+3 long and 2+2 short setae. Tergites 3–20 with paramedian sulci, without dark subcutaneous pigment, posterior margin of sternite 21 straight, coxopleuron with 26 pores. Ultimate leg tibia with 7–8, tarsus with 2–4 saw teeth. Tarsi of most legs more or less divided.
China.
Cryptops songi would appear to be quite distinct from other Chinese species of the genus. It is clearly related to Cryptops nigropictus, Cryptops decoratus and Cryptops melanotypus but further data are required to establish its exact relationship to them.
http://species-id.net/wiki/Cryptops_stabilis
Figs 86–91ANSP Holotype labelled Type 9921/lot 1398. Cryptops stabilis Ch. Malabar XII-1920 in a small unlabelled micro vial in a larger vial in which there was a second specimen placed in a second microvial and labelled Cryptops stabilis second specimen by J. G. E. L. The two are certainly conspecific.
NB. Chamberlin stated that the type from Goenoeng (=Gunung?) Malabar, Java was the only specimen seen.
Length 11 mm. Without dark subcuticular pigment. Cephalic plate and all tergites without paramedian sutures or sulci. Each side of coxosternite with 2 large and 2–3 smaller submarginal setae. Poison calyx oval. Coxopleural pores large, pore field occupying anterior 60% of coxopleuron. Ultimate leg prefemur with medial longitudinal glabrous strip and with 6 tibial and 3 tarsal saw teeth. Legs 1–20 with minute pretarsal accessory spurs.
(
Length 11 mm. Cephalic plate and tergite 1 orange, trunk light brown with darker subcuticular pigment (head and first and last tergites yellow or orange, the remaining portion of body dark).
Two basal antennal articles with long setae, third with long and many short, fourth and subsequent with dense short setae and irregular basal whorl of long setae. Cephalic plate without sutures. Tergite 1 without sutures and overlying cephalic plate. Anterior margin of forcipular coxosternite curved (widely convex) with 2 large and 2 small submarginal setae on each side (Fig. 86). Poison gland calyx oval.
Tergites apparently without paramedian sutures or sulci in either specimen (median sulci absent). Lateral crescentic sulci on tergites 3–18. Sternites with cruciform sulci. Transverse skeletal thickening not observable. Sternite 21 in spm. 2 with posterior margin weakly convex.
Coxopleuron with 14+16 pores of varying size (Fig. 87), the pore field, which is without setae, occupying the anterior 64% of coxopleuron and 54% in spm 2. Two setae between the pore field and the posterior margin on which there are 4 large and 2 small setae. Prefemur of ultimate leg (Fig. 88) with strong setae (Fig. 89) of moderate length on prefemur, some long ventral setae with an elongated glabrous strip on medial face. Femur with some strong setae, tibia and tarsus 1 and 2 with fine setae. Tibia with 6, tarsus with 3 saw teeth (Fig. 90). Legs 1–19 with undivided tarsi and fine setae. Leg 20 with dense setae ventrally on distal half of prefemur and ventrally on femur and tibia. Pretarsi of legs 1–20 long, with minute accessory spurs (Fig. 91).
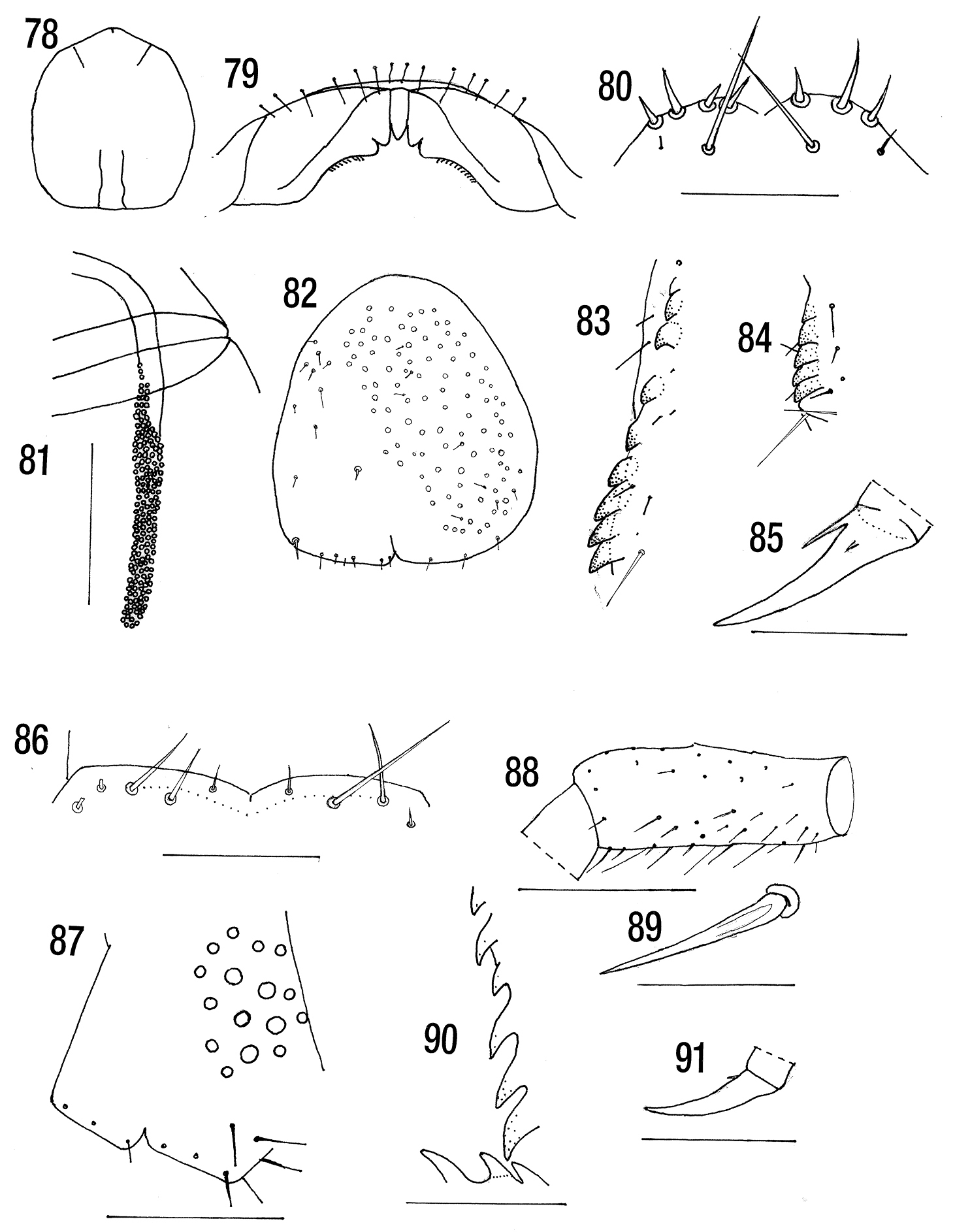
Cryptops parisi parisi. 78 Cephalic plate (after
Status of the species of the Cryptops hortensis group of the Old World.
| Species | Authority | Distribution | Status |
|---|---|---|---|
| Cryptops basilewskyi | Matic & Darabantu, 1977 | St. Helena | = Cryptops nigropictus Syn. n. |
| Cryptops brignolii | Matic, 1977 | Turkey | Valid pro tem |
| Cryptops campestris | Attems, 1953 | Rwanda & DRC | Nomen dubium |
| Cryptops compositus | Chamberlin, 1952 | Turkey | Nomen dubium |
| Cryptops covertus | Chamberlin, 1951 | Angola | Valid |
| Cryptops daszaki | Lewis, 2002 | Mauritius | Valid |
| Cryptops decoratus | Lawrence, 1960 | Madagascar | Valid |
| Cryptops decoratus | sensu Lewis, 2002, 2007 | Mauritius, Seychelles | = Cryptops melanotypus Syn. n. |
| Cryptops h. hortensis | (Donovan, 1810) | Europe, Central Asia, widely introduced | Valid |
| Cryptops hortensis atlantis | Pocock, 1891 | Madeira (?Azores, Canaries) | Cryptops atlantis Stat. n. |
| Cryptops inermipes | Pocock, 1888 | Christmas I. | Valid |
| Cryptops kalobensis | Goffinet, 1971 | Democratic Republic Congo | = Cryptops nigropictus Syn. n. |
| Cryptops lobatus | Verhoeff, 1931 | France, Italy | Valid |
| Cryptops mauritianus | Verhoeff, 1939 | Mauritius | Valid |
| Cryptops medius | Verhoeff, 1901 | Greece | Nomen dubium |
| Cryptops melanotypus | Chamberlin, 1941 | Philippines, Mauritius, Seychelles | Valid |
| Cryptops nigropictus | Takakuwa, 1936 | Afrotropical & Oriental regions | Valid |
| Cryptops niloticus | Lewis, 1967 | Afrotropical region | = Cryptops nigropictus Syn. n. |
| Cryptops orthus | Chamberlin, 1951 | Angola | Nomen dubium |
| Cryptops parisi parisi | Brolemann, 1920 | Europe, Turkey, Canada (introduced) | Valid |
| Cryptops parisi cottarellii | Matic, 1977 | Turkey | = Cryptops parisi parisi Syn. n. |
| Cryptops parisi rhenanus | Verhoeff, 1931 | Germany | = Cryptops parisi parisi Syn. n. |
| Cryptops parisi sebini | Verhoeff, 1934 | Italy | Nomen dubium. |
| Cryptops penicillatus | Lawrence, 1960 | Madagascar | = Cryptops decoratus Syn. n. |
| Cryptops philammus gabonensis | Demange, 1968 | Gabon | Nomen dubium |
| Cryptops songi | Song et al., 2010 | China | Valid |
| Cryptops stabilis | Chamberlin, 1944 | Indonesia (Java) | Valid |
Indonesia (Java).
If absence of tergite paramedian sutures and sulci is disregarded (possible juvenile character) then the species is close to Cryptops melanotypus.
Provisional key to Cryptops s. str. species of the hortensis group
| NB. |
|
|
| 1 | Tibia and tarsus of ultimate leg without saw teeth. Christmas Island | Cryptops inermipes Pocock, 1888 |
| – | Tibia and tarsus of ultimate leg with saw teeth | 2 |
| 2 | With the anterior edge of the forcipular coxosternite strongly bilobed. Each lobe with five short marginal setae (Fig. 66). France, Italy | Cryptops lobatus Verhoeff, 1931 |
| – | Anterior edge of the forcipular coxosternite not strongly lobed (at most as in Cryptops covertus) (Fig. 17) | 3 |
| 3 | Prefemur of ultimate leg with glabrous longitudinal ventral groove | 4 |
| – | Prefemur of ultimate leg without glabrous longitudinal ventral groove | 5 |
| 4 | Coxosternal setae submarginal (Fig. 39), tibial saw teeth 5–8, tarsal 2–4. Europe and Atlantic Islands, western Central Asia. Widely introduced elsewhere | Cryptops hortensis (Donovan, 1810) |
| – | Coxosternal setae fine, marginal (Fig. 46), tibial saw teeth 15–17, tarsal 5–6. Madeira, ?Azores and Canary Is | Cryptops atlantis Pocock, 1891 |
| 5 | Cephalic plate with paramedian sutures occupying posterior 30% (Fig. 78), labrum usually tripartite (Fig. 79), with 50 or more small coxal pores. Europe and SW Asia (Turkey) | Cryptops parisi Brolemann, 1920 |
| – | Cephalic plate without (Fig. 1) or with very short posterior paramedian sutures, labrum unipartite, coxopleuron with maximum of 26 (rarely to 31) pores | 6 |
| NB. 1 | Up to 50 pores in Cryptops. cf. hortensis a high altitude (1, 000–1, 300 m) population described from Karnten, Austria, by |
|
| NB. 2 | The inadequately described Cryptops brignolii near here. | |
| 6 | Anterior edge of forcipular coxosternite bilobed with 6+6 stout marginal setae (Fig. 17) and about 25 coxopleural pores. Angola | Cryptops covertus Chamberlin, 1951 |
| – | Anterior edge of forcipular coxosternite not markedly bilobed and with fine setae | 7 |
| 7 | 20–32 mm, with 33–65 coxopleural pores, pretarsal accessory spurs minute (Fig. 70), 3+4–5+5 long submarginal coxosternal setae (Fig. 68). Mauritius | Cryptops mauritianus Verhoeff, 1939 |
| – | Maximum length 20 mm, with 7–25 (rarely 31 in Sudanese Cryptops nigropictus) pretarsal accessory spurs short to long (except in Cryptops stabilis where they are also minute) | 8 |
| 8 | Tergites without paramedian sutures or sulci, pretarsal accessory spurs minute. Length 11 mm. Java (may be an immature specimen) | Cryptops stabilis Chamberlin, 1944 |
| – | Tergites with paramedian sutures (may be incomplete), pretarsal accessory spurs may be short but not minute | 9 |
| 9 | Pretarsi of ambulatory legs each with a single long accessory pretarsal spur | 10 |
| – | Pretarsi of ambulatory legs with two accessory spurs, the second sometimes very small and easily overlooked | 11 |
| Cryptops songi Song et al., 2010 a species with short cephalic posterior paramedian sutures, without subcutaneous pigment and with 26+26 coxopleural pores, China (near here). | ||
| (The inadequately described Cryptops orthus Chamberlin, 1951, Angola, and Cryptops campestris Attems, 1953, Rwanda and Democratic Republic of Congo also near here). | ||
| 10 | Forcipular coxosternite with 2+3 fine marginal setae. Ultimate leg tibia with 3 saw teeth, tarsus with 2. Ambulatory legs with single long pretarsal accessory spurs (Fig. 27). 7.5 mm. Without dark subcuticular pigment. Mauritius (Ile aux Aigrettes) | Cryptops daszki Lewis, 2002 |
| – | Forcipular coxosternite generally with 2+2 long submarginal setae and several shorter setae. Ultimate leg tibia with 4–8 saw teeth, tarsus with (2)3–4. With dark subcutaneus pigment. St. Helena, Sudan, Eritrea, Somalia, ?Yemen, Democratic Republic of Congo, Rodrigues, Taiwan | Cryptops nigropictus Takakuwa, 1936 |
| 11 | With 6–9 coxopleural pores, 5–9 setae between pore field and coxopleural margin, ultimate prefemur without medial longitudinal glabrous area. Madagascar | Cryptops decoratus Lawrence, 1960 |
| – | With 7–16 coxopleural pores, 1–3(4) setae between pore field and coxopleural margin, ultimate prefemur with broad medial longitudinal glabrous area. Philippines, Mauritius, Seychelles | Cryptops melanotypus Chamberlin, 1941 |
The separation of those Cryptops s. str. species lacking an anterior transverse suture on tergite 1 into two groups dependant on the presence (doriae group) or absence (hortensis group) of a saw tooth or teeth on the ultimate femur may prove to be artificial but is adopted as a convenient way of dealing with of this group of species pro tem. The character has been overlooked by some workers in the past and species that appear from their description to belong the hortensis group in fact belong to the doriae group (see Introduction). Evidence to suggest that this separation may, however, be a natural one, is provided by the fact that no species in the doriae group have been recorded from Europe and no members of the hortensis group have been recorded as naturally occurring in Australasia or the Afrotemperate region.
Ten of the species keyed out here are quite distinct. However, the status of Cryptops decoratus, Cryptops melanotypus and Cryptops nigropictus, and the species that have here been synonymised with them and of Cryptops songi is uncertain. These species would appear to be closely related. Cryptops nigropictus has been separated from Cryptops decoratus and Cryptops melanotypus on the basis of the nature of the pretarsal spur(s): single and long in Cryptops nigropictus, two and generally shorter in the other two “species”.
Cryptops niloticus fromSudan, Yemen, Rodrigues, Somalia and possibly Eritrea, Cryptops kalobensis from Democratic Republic of Congo and Cryptops basilewskyi from St Helena are synonymised under Cryptops nigropictus
itself recorded from China, Taiwan and the Ryuku Islands. It is thus a
very widely distributed species. The Sudanese populations differ from
others in that some specimens are larger; have more coxosternal pores
and more submarginal coxosternital setae. A similar situation was
reported by
The synonymising of Cryptops decoratus sensu (
I am greatly indebted to the following for the loan of material: Don Azuma (ANSP), Jan Beccaloni (BMNH), Jean-Paul Mauriès (MNHN), Jonathan Coddington, Scott F. Larcher, Gary F. Hevel and David G. Furth (NMNH), Jürgen Gruber and Verena Stagl (NMW) and Rudy Joqué (MRAC). My thanks are also due to Dennis Parsons and staff of the former Somerset County Museum, now part of the Somerset Heritage Service, for providing excellent working conditions in that Museum. Thanks also to Roger Lewis for processing the figures, to an anonymous reviewer for very detailed and constructive comments on the manuscript and to Pavel Stoev for his help and advice.