





(C) 2011 Karin Voigtländer. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Knowledge of habitat preferences of myriapod species is essential for their optimal use in bioindication and biological soil assessment. To increase our knowledge a sampling project was started in Saxony-Anhalt (Eastern Germany) more than 20 years ago. The narrowly defined geographic area was selected for the study because the fauna of this Central European German region represents to a high degree the whole of the Central European fauna. The millipede fauna at 175 sites belonging to 12 biotope types was sampled by pitfall trapping and identified to species. According to plant associations, these sites could be divided into four groups differing in moisture and into two groups differing in the level of vegetation cover. Considering the distribution pattern of 28 selected millipede species in each of these groups (weighted percentage), ecological categories of these species could roughly be distinguished. “Main Preferences” separated open land or woodland species and hygrobiont or xerobiont species. The number (and quality) of the occupied biotope types allowed assessment as stenotopic or eurytopic species (“General Preferences”). If available, a “Special Preference” for a preferred biotope type was recognised. For several species, the results are compared with those in the literature.
Diplopoda, woodland species, open land species, biotope type, preference, hygrobiont, xerobiont, eurytopic, stenotopic
In the last 10 to 15 years the protection of structure and function of soil biocoenoses has become more and more a focus of public and governmental interest, and methods for assessing biological soil quality are in demand. Diplopods as saprophagous animals are an essential component of soil fauna. Their role as bioindicators is indisputable. However, before diplopods can be used as optimal indicators for site characterisation, a better knowledge of their environmental requirements will be needed. Laboratory autecological experiments are unfortunately rare (
Ecological preferences of animals are mostly deduced from characteristics of the habitat in which these animals are living (e.g. sandy/loamy soils, under grasses or woodlands, in open soils, etc.). More detailed information is available from the description of plant associations and their characteristics. To increase our knowledge of diplopod habitat selection, the millipede fauna of different biotope types defined by plant sociological studies in the German federal state of Saxony-Anhalt (Central Europe) was investigated and compared with literature data. The narrowly defined geographic area was selected for the study because the fauna of this Central European German region represents to a high degree the Central European fauna as a whole.
Methods and sites MaterialOver the past 20 years the Senckenberg Museum of Natural History Görlitz (SMNG) has been studying the soil fauna at a variety of sites spread out over the whole area of Saxony-Anhalt (e.g.
Descriptions of sampling sites are based on defined plant associations (= biotope types; see Table 1). Moisture and temperature were chosen as the most important habitat factors determining distribution of myriapods (
Description of sampling sites.
| Biotope type | Plant association | Number of studied sites | Site characters | |
|---|---|---|---|---|
 |
bogs and swamps | Sphagnetalia magellanici Scheuchzerietalia palustris | 14 | very wet, single-level vegetation cover |
 |
fresh meadows and pastures | Molinio-Arrhenatheretea, Dauco-Arrhenatheretum (fresh formation), Trisetetum flavescentis, Angelico-Cirsietum oleracei | 8 | wet to humid, single-level vegetation cover |
 |
brook- or riversides | Alnus- or Alnus-Fraxinus- and Salix-associations | 5 | wet to humid, multi-level vegetation cover |
 |
flood plains and swamp forests | Alnetum, Betuletum, Salicetum, Ledo palustris-Pinetum | 19 | very wet, multi-level vegetation cover |
 |
deciduous and deciduous mixed forests |
Fagetalia, Querco-Carpinetum | 25 | wet to humid, multi-level vegetation cover |
 |
thermophilic woods | Quercetalia pubescenti-petraeae, Tilio-Carpinetum on sandy-loamy soils | 6 | dry, multi-level vegetation cover |
 |
coniferous forests | Pinetum | 14 | dry, multi-level vegetation cover |
 |
dwarf-shrub heaths (DSH) | – subatlantical DSH (Genisto pilosae-Callunetum) on nutrient-poor, sandy soil (podsol) | 9 | very dry, single-level vegetation cover |
| – subalpine DSH (Pulsatillo- Nardetum), on granite gravel (Euphorbio-Callunetum, Harz mountain) | ||||
 |
xeric shrub societies | Berberidion with the association Ligustro-Prunetum spinosae and pioneer forests tending towards the association Potentillo albae-Quercetum petraeae on loamy-mary soils | 6 | dry, multi-level vegetation cover |
 |
xeric and mesoxeric meadows | – Corynephorion canescentis with Spergulo morisonii-association | 54 | very dry, single-level vegetation cover |
| – Corynephoretum canescentris on dry and warm, nutrient-poor inland dunes with loose Pleistocene sand | ||||
| – Armerion elongatae on sandy, dry soils with a more or less multi-level sod | ||||
| – Teucrio-Seslerietum | ||||
| – Festuco-Brachypodietum | ||||
| – Mesobrometum | ||||
| – Festuco-Stipetum | ||||
 |
special structures |
– semi-natural and anthropogenic vegetation-poor sites (clear cutting, granite debris) | 5 | dry, single-level vegetation cover |
 |
fields and fallows on different soils | 8 | dry, single-level vegetation cover | |
The ecological classification of species was carried out as follows (Fig. 1):
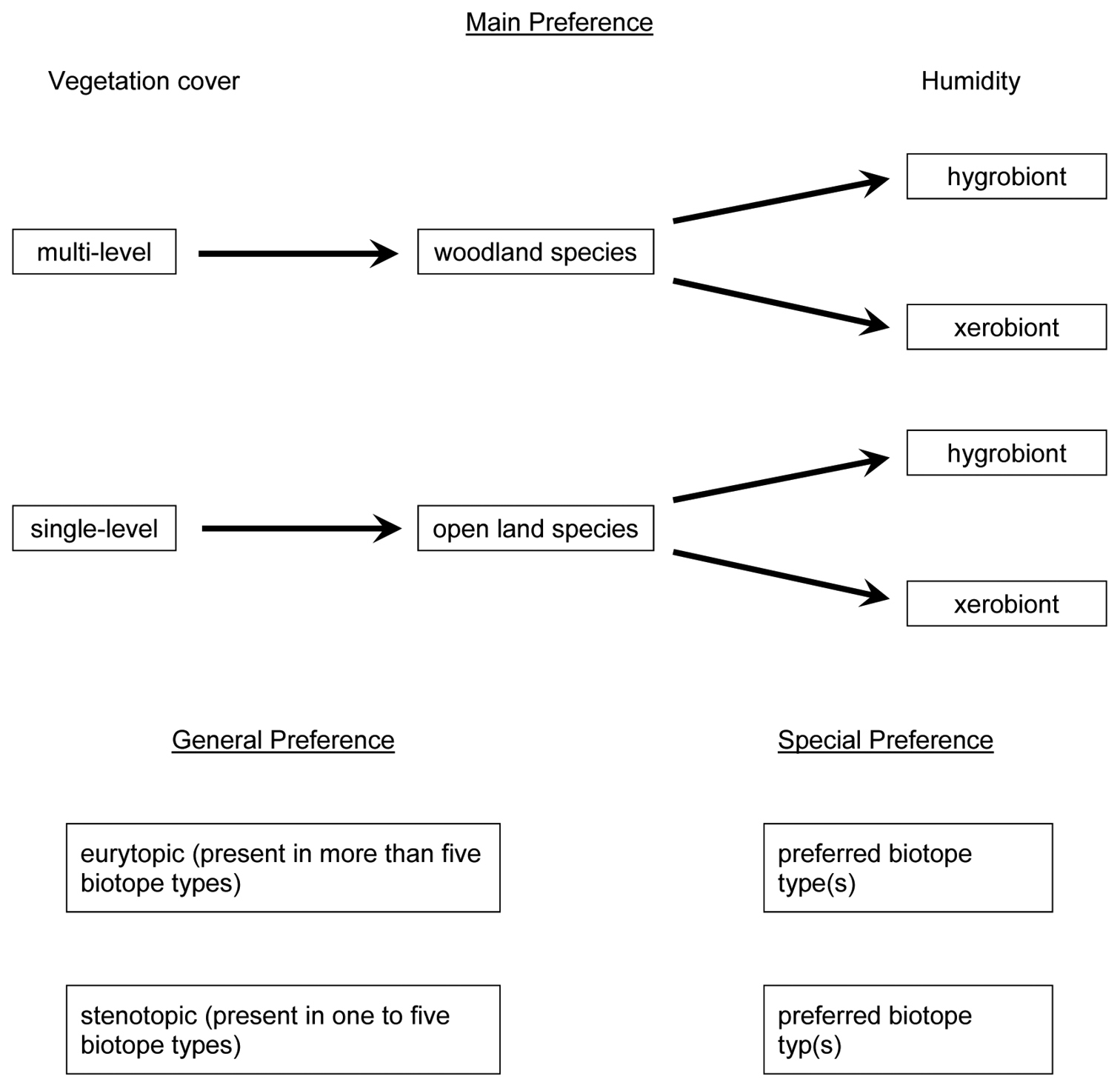
Classification of preferences.
For “General Preferences” (distribution over 12 biotope types) a species was said to be eurytopic or stenotopic. Occurrence in one to five (similar) biotope types defined a stenotopic species; occurrence in six or more biotope types, a eurytopic species; and occurrences in fewer than six but very different biotope types, a eurytopic species.
“Main Preferences” were deduced from the species distribution along a moisture and vegetation gradient and from an assessment of a species as woodland vs open land and hygrobiont vs xerobiont.
“Special Preferences” refers to occurrence in special biotope types (e.g. mesoxeric meadow, mixed forest).
CalculationThe Main Preference of millipede species at particular site types was determined from their “presence”, which was calculated using the number of different sites of the same biotope-type (within a greater area) at which a certain species occurs:
P = presence
Si = the number of sites in which a particular species occurs
Xi = the total number of sites
As different site numbers of each biotope type were investigated, a weighting had to be included according to the following formula:
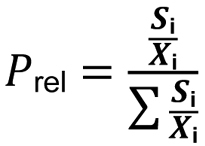
On the basis of distribution over the 12 biotope types, 20 species can be characterised as eurytopic and eight species as stenotopic (Figs 11, 12, 14, 20, 22, 24, 25 and 27), with stenotopy especially high in open land species. Although Julus scanicus, Megaphyllum projectum kochi and Brachydesmus superus occur only in five biotope types (Figs 10, 19 and 28) these species are assessed as eurytopic, because the site characteristics differ widely.
Main preferencesTable 2 summarises species distribution along the moisture and vegetation cover gradients of the 175 investigated sites. Eight inhabitant groups are recognised:
Species distribution (groups of Main Preference) along the moisture and vegetation cover gradients of 175 investigated sites using Prel (see above).
| Degree of humidity | Vegetation cover | Family | |||||
|---|---|---|---|---|---|---|---|
| very wet | wet to humid | dry | very dry | single-level | multi-level | ||
| Group I Inhabitants of biotope types with multi-level vegetation cover and different moisture (woodland species) | |||||||
| Glomeris undulata var. conspersa C. L. Koch, 1847 | – | 38.67 | 28.67 | 32.66 | 36.84 | 63.16 | Glomeridae |
| Glomeris marginata (Villers, 1789) | 8.02 | 34.82 | 19.36 | 37.80 | 46.52 | 53.48 | Glomeridae |
| Glomeris hexasticha Brandt, 1833 | 19.70 | 23.96 | 25.38 | 30.96 | 38.71 | 61.29 | Glomeridae |
| Proteroiulus fuscus (Am Stein, 1857) | 30.19 | 19.66 | 30.38 | 19.77 | 34.43 | 65.57 | Blaniulidae |
| Leptoiulus proximus (Němec, 1896) | 16.75 | 36.37 | 33.71 | 13.16 | 27.27 | 72.73 | Julidae |
| Julus scandinavius Latzel, 1884 | 25.17 | 31.80 | 20.26 | 22.77 | 33.66 | 66.34 | Julidae |
| Group II Inhabitants of very wet biotope types with multi-level vegetation cover (hygrobiont woodland species) | |||||||
| Polyzonium germanicum Brandt, 1831 | 88.56 | 4.52 | 4.19 | 2.73 | 11.69 | 88.31 | Polyzoniidae |
| Craspedosoma rawlinsii (Leach, 1815) | 38.05 | 34.78 | 17.73 | 9.44 | 30.84 | 69.16 | Craspedosomatidae |
| Polydesmus denticulatus C. L. Koch, 1847 | 46.44 | 33.61 | 14.54 | 5.41 | 20.00 | 80.00 | Polydesmidae |
| Xestoiulus laeticollis (Porat, 1889) | 94.53 | 5.47 | – | – | 20.00 | 80.00 | Julidae |
| Julus scanicus Lohmander, 1925 | 65.69 | 19.02 | – | 15.29 | 31.03 | 68.97 | Julidae |
| Group III Inhabitants of wet to humid biotope types with multi-level vegetation cover (hygrobiont woodland species) | |||||||
| Polydesmus angustus (Latzel, 1884) | 3.04 | 50.18 | 24.48 | 22.30 | 30.00 | 70.00 | Polydesmidae |
| Nemasoma varicorne C. L. Koch, 1844 | 32.63 | 47.22 | 8.75 | 11.39 | 15.79 | 84.21 | Nemasomatidae |
| Allajulus nitidus (Verhoeff, 1891) | 11.58 | 45.27 | 27.97 | 15.17 | 34.18 | 65.82 | Julidae |
| Tachypodoiulus niger (Leach, 1815) | 2.67 | 55.63 | 19.33 | 22.37 | 31.49 | 68.51 | Julidae |
| Unciger foetidus (C. L. Koch, 1838) | 11.32 | 55.71 | 15.19 | 17.79 | 29.03 | 70.97 | Julidae |
| Group IV Inhabitants of very dry biotope types and multi-level vegetation cover (xerobiont woodland species) | |||||||
| Leptoiulus belgicus (Latzel, 1884) | 3.70 | 28.88 | 26.77 | 40.65 | 45.32 | 54.68 | Julidae |
| Group V Inhabitants of biotope types with single-level vegetation cover and different moisture (open land species) | |||||||
| Megaphyllum projectum kochi (Verhoeff, 1907) | 41.37 | 7.19 | 6.66 | 44.78 | 54.50 | 45.50 | Julidae |
| Group VI Inhabitants of very dry and dry biotope types with single-level vegetation cover (xerobiont open land species) | |||||||
| Polydesmus inconstans Latzel, 1884 | 10.33 | 24.67 | 31.18 | 33.82 | 68.72 | 31.28 | Polydesmidae |
| Melogona voigtii (Verhoeff, 1899) | 14.12 | – | 34.10 | 51.78 | 56.76 | 43.24 | Chordeumatidae |
| Choneiulus palmatus (Němec, 1895) | 6.30 | 5.47 | 35.47 | 52.77 | 89.61 | 10.39 | Blaniulidae |
| Ophyiulus pilosus (Newport, 1842) | 25.13 | 10.91 | 60.67 | 3.29 | 80.00 | 20.00 | Julidae |
| Kryphioiulus occultus (C. L. Koch, 1847) | – | – | 18.00 | 82.00 | 100.00 | – | Julidae |
| Cylindroiulus caeruleocintus (Wood, 1864) | 3.66 | 28.59 | 29.44 | 38.32 | 60.90 | 39.10 | Julidae |
| Megaphyllum unilineatum (C. L. Koch, 1838) | 0.00 | 4.00 | 33.33 | 62.67 | 96.33 | 3.67 | Julidae |
| Ommatoiulus sabulosus (Linnaeus, 1758) | 14.93 | 12.97 | 24.04 | 48.05 | 71.68 | 28.32 | Julidae |
| Group VII Inhabitants of very wet biotope types and different vegetation cover (hygrobiont) | |||||||
| Brachydesmus superus Latzel, 1884 | 48.03 | 11.92 | 22.09 | 17.97 | 48.39 | 51.61 | Polydesmidae |
| Group VIII No influence of moisture or vegetation cover | |||||||
| Mycogona germanica (Verhoeff, 1892) | 17.41 | 30.24 | 28.03 | 24.32 | 51.22 | 48.78 | Chordeumatidae |
Woodland species: Of the 17 species characterised as woodland species, 10 species are hygrobiont (groups II and III) with a preference for very wet biotope types (group II, five species) or wet to humid biotope types (group III, five species). One woodland species, Leptoiulus belgicus, is xerobiont (group IV). On the distribution of six species (group I), the moisture level of the biotope types has no apparent influence.
Open land species: Only nine species can be characterised as open land species (groups VI and V), and most of these are xerobiont. Only the occurrence of Megaphyllum projectum kochi is not apparently influenced by the moisture level of the biotope type. Eight species show a clear preference for a combination of very dry or dry biotope types with single-level vegetation cover. Only Polydesmus inconstans does not differ between these two moisture categories. One species (Ophyiulus pilosus) prefers dry, the other six species prefer very dry biotope types (xeric and mesoxeric meadows).
Uncharacterised species: Brachydesmus superus and Mycogona germanica do not fit into the scheme of woodland or open land species. Brachydesmus superus is clearly hygrobiont (group VII), whilefor Mycogona germanica no influence of moisture or vegetation cover could be observed (group VIII).
Special preferencesThe distribution of each investigated species over the biotope types can be seen in Figs 2–29. Depending on the level of occurrence, Special Preferences can be recognised.
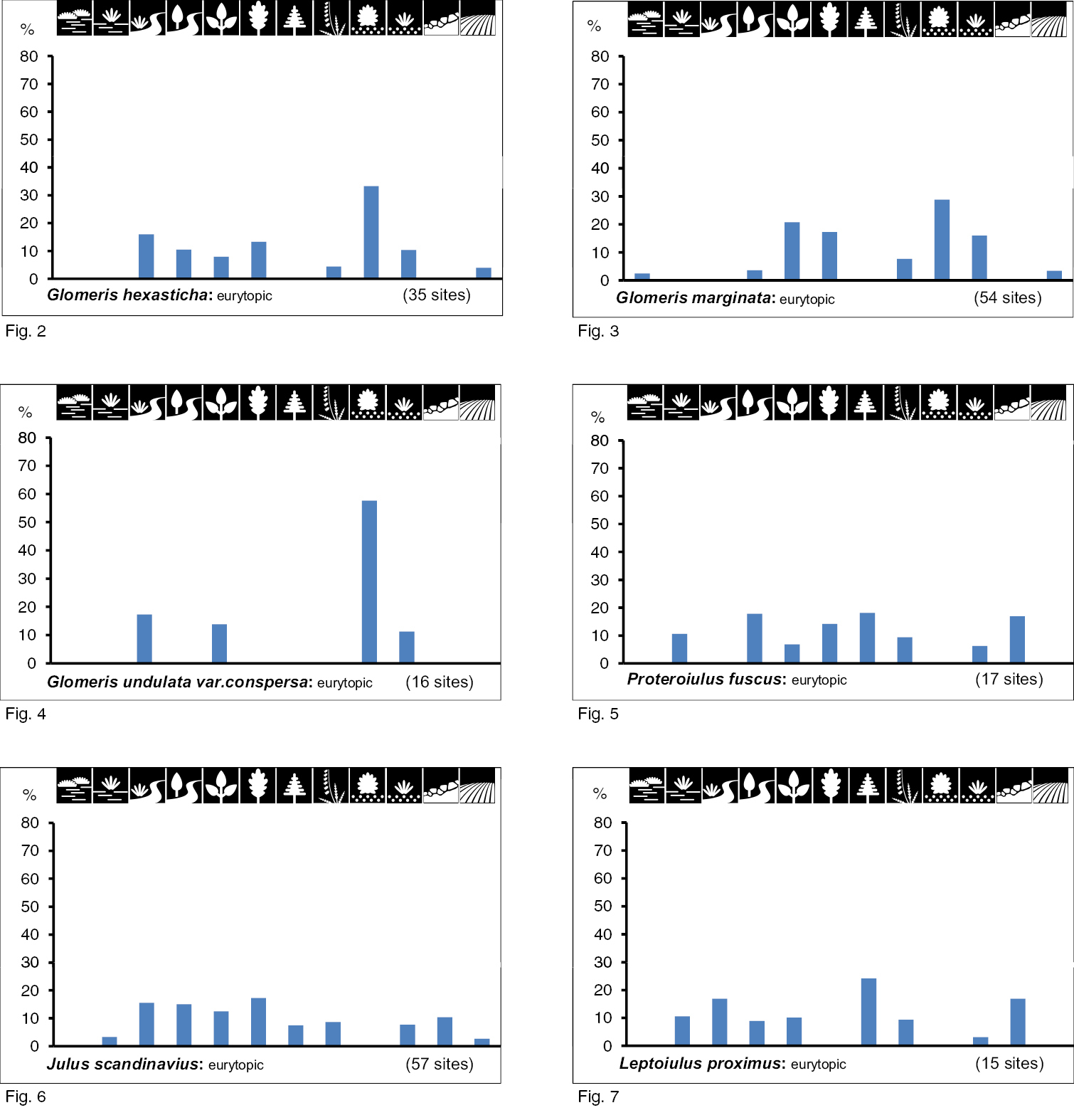
Occurrence of the species of group I (inhabitants of biotope types with multi-level vegetation cover and different moisture) in the 12 biotope types investigated. Numbers of positive samples are given in brackets.
Occurrence of the species of group II (inhabitants of very wet biotope types with multi-level vegetation cover) in the 12 biotope types investigated. Numbers of positive samples are given in brackets.
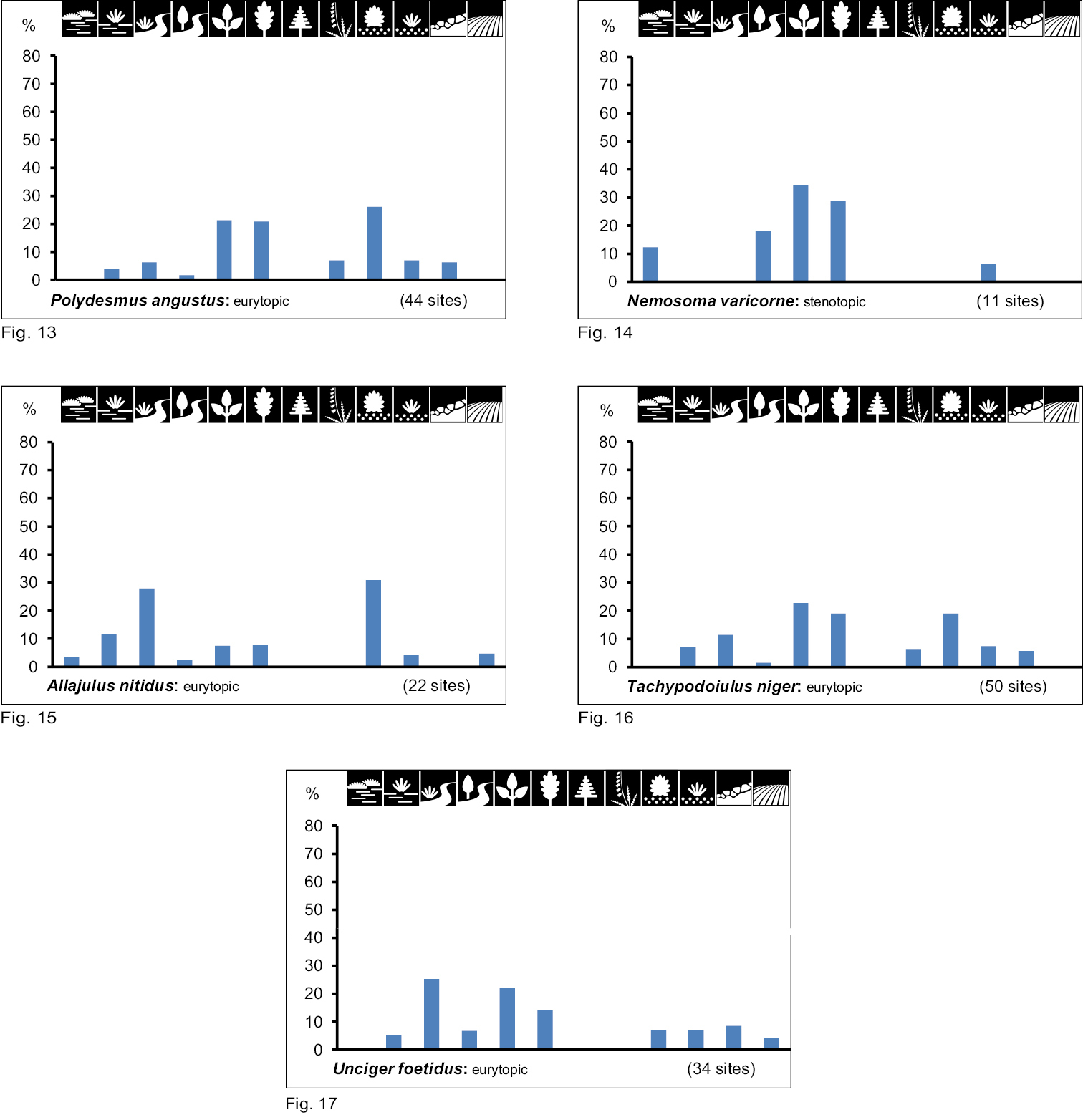
Occurrence of the species of group III (inhabitants of wet to humid biotope types with multi-level vegetation cover) in the 12 biotope types investigated. Numbers of positive samples are given in brackets.
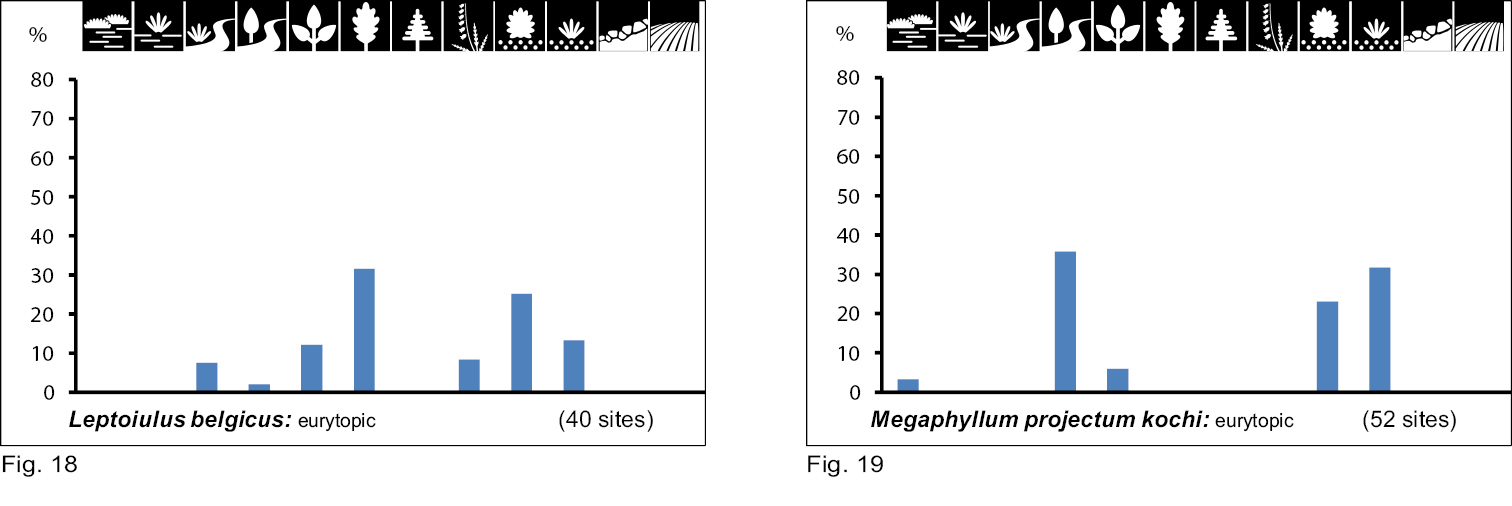
Occurrence of the species of group IV and V in the 12 biotope types investigated. Numbers of positive samples are given in brackets.
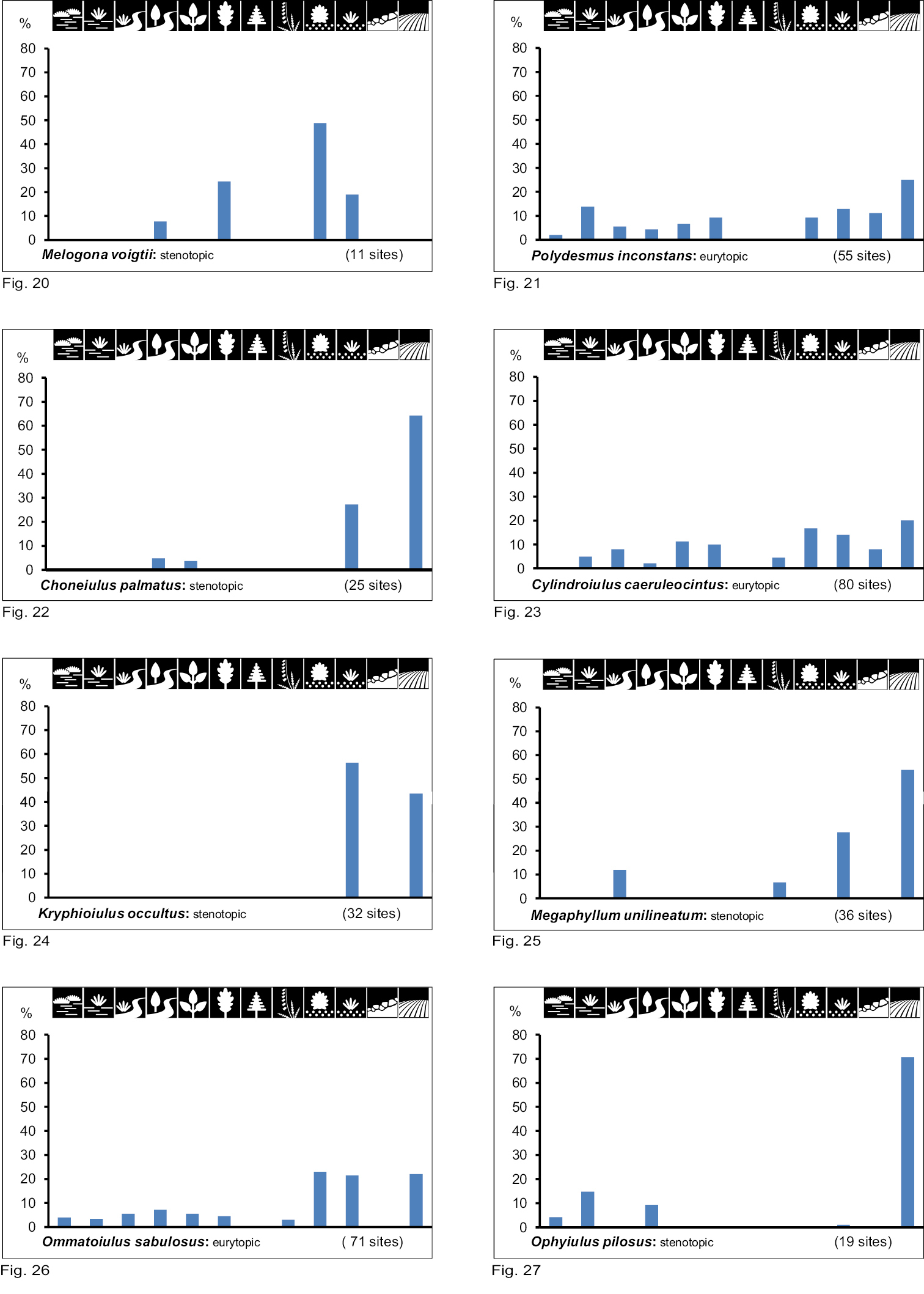
Occurrence of the species of group VI (inhabitants of very dry and dry biotope types with single-level vegetation cover) in 12 biotope types investigated. Number of positive samples are given in brackets.
The biotope type selections of certain species are discussed below.
Combination of preferencesAn assessment of the autecology of the species investigated can be derived from a combination of the three preference types (General, Main and Special). An overview is given in Table 3.
Ecological characterisation of selected Central European diplopods in Saxony-Anhalt.
| Species | General Preference | Main Preference | Special Preference |
|---|---|---|---|
| Glomeris undulata var. conspersa | eurytopic | woodland species | with preference for xeric shrub societies |
| Glomeris hexasticha | eurytopic | woodland species | with preference for xeric shrub societies |
| Glomeris marginata | eurytopic | woodland species | without clear preferences |
| Julus scandinavius | eurytopic | woodland species | without clear preferences |
| Leptoiulus proximus | eurytopic | woodland species | without clear preferences |
| Proteroiulus fuscus | eurytopic | woodland species | without clear preferences |
| Allajulus nitidus | eurytopic | hygrobiont woodland species | without clear preferences |
| Craspedosoma rawlinsii | eurytopic | hygrobiont woodland species | with preference for floodplains, riparian woody sites and deciduous woods |
| Polydesmus angustus | eurytopic | hygrobiont woodland species | without clear preferences |
| Polydesmus denticulatus | eurytopic | hygrobiont woodland species | without clear preferences |
| Polyzonium germanicum | stenotopic | hygrobiont woodland species | with preference for floodplains and swamp forests |
| Julus scanicus | eurytopic | hygrobiont woodland species | with preference for floodplains and swamp forests |
| Tachypodoiulus niger | eurytopic | hygrobiont woodland species | without clear preferences |
| Unciger foetidus | eurytopic | hygrobiont woodland species | without clear preferences |
| Xestoiulus laeticollis | stenotopic | hygrobiont woodland species | with preference for floodplains and swamp forests |
| Nemosoma varicorne | stenotopic | hygrobiont woodland species | without clear preferences |
| Brachydesmus superus | eurytopic | hygrobiont | with preference for floodplains and swamp forests, often synanthropic |
| Leptoiulus belgicus | eurytopic | xerobiont woodland species | with preference for thermophilous oak woods and xeric shrub societies |
| Megaphyllum projectum kochi | eurytopic | open land species | without clear preferences |
| Cylindroiulus caeruleocintus | eurytopic | xerobiont open land species | with preference for fields and fallows |
| Ommatoiulus sabulosus | eurytopic | xerobiont open land species | without clear preferences |
| Polydesmus inconstans | eurytopic | xerobiont open land species | without clear preferences |
| Mycogona germanica | eurytopic | – | without clear preferences |
| Choneiulus palmatus | stenotopic | xerobiont open land species | with preference for xeric/mesoxeric meadows, fields and fallows |
| Kryphioiulus occultus | stenotopic | xerobiont open land species | with preference for xeric/mesoxeric meadows, fields and fallows |
| Megaphyllum unilineatum | stenotopic | xerobiont open land species | with preference for xeric/mesoxeric meadows, fields and fallows |
| Melogona voigtii | stenotopic | xerobiont open land species | with preference for xeric shrub societies |
| Ophyiulus pilosus | stenotopic | xerobiont open land species | with preference for fields and fallows, often synanthropic |
Species belonging to group I are woodland species. They are eurytopic, mostly without special preferences for a biotope type. Occurrence in eight to 10 different types is the rule (see Figs 2 to 7). Their occurrence seems to be influenced by the reduction in insolation and temperature (closed tree or shrub layer), and less by moisture.
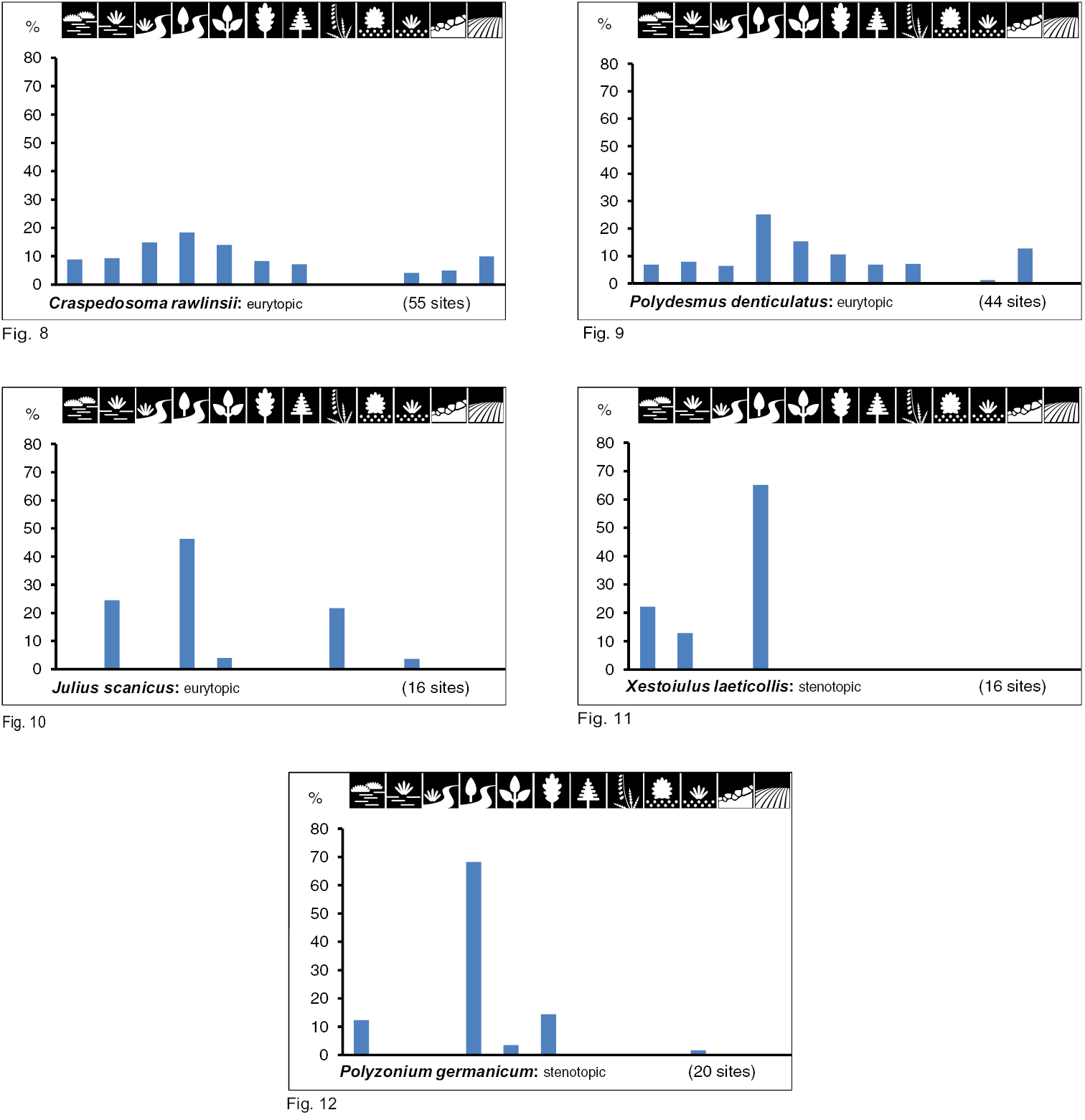
Group II includes inhabitants of very wet biotope types with multi-level vegetation cover. They can be classified as hygrobiont woodland species. Biotopes with very wet soils (sometimes inundated) under a multi-level vegetation cover were preferred by 5 species, among which Craspedosoma rawlinsii and Polydesmus denticulatus (Figs 8 and 9) have a very broad biotope spectrum in total (10 biotope types; eurytopic). Polyzonium germanicum, Julus scanicus and especially Xestoiulus laeticollis occur in fewer types (Figs 10 to 12). With the exception of Julus scanicus, which lives in five very different biotope types, they can be characterised as stenotopic. However, all five species have in common a preference for floodplains and swamp forests (in Figs 10 to 12; the fourth biotope type).
Group III consists of five species which inhabit wet to humid biotope types with multi-level vegetation cover. They can be characterised as hygrobiont woodland species. With the exception of Nemasoma varicorne (stenotopic, Fig. 14) all these species may inhabit seven to nine different biotope types (eurytopic; Figs 13, 15–17).
To group IVbelongs the one xerobiont woodland species, Leptoiulus belgicus. It inhabits seven different biotope types (Fig. 18), which results in its characterisation as eurytopic.
Group Vare open land species, among which Megaphyllum projectum kochi is the only species of those investigated. It occurs in five biotope types (Fig. 19) which are very different in vegetation cover and moisture level, and is therefore assessed as eurytopic.
Group VI consists of xerobiont open land species. Eight species show a clear preference for a combination of very dry/ dry biotope types with single-level vegetation cover (Table 2). Within this species group we can also differentiate between species with a wide biotope spectrum (eurytopic), namely Polydesmus inconstans (Fig. 21), Cylindroiulus caeruleocinctus (Fig. 23) and Ommatoiulus sabulosus (Fig. 26), and others with high specialisation for only a few biotope types (stenotopic), namely Ophyiulus pilosus (Fig. 27), Megaphyllum unilineatum (Fig. 25), Choneiulus palmatus (Fig. 22), Melogona voigtii (Fig. 20), and Kryphioiulus occultus (Fig. 24).
Brachydesmus superus occurs in very wet biotope types with variable vegetation cover (group VII), and therefore cannot be classified as a woodland or open land species. The species is eurytopic, but with a preference for very wet and humid places (Table 2, Fig. 28).
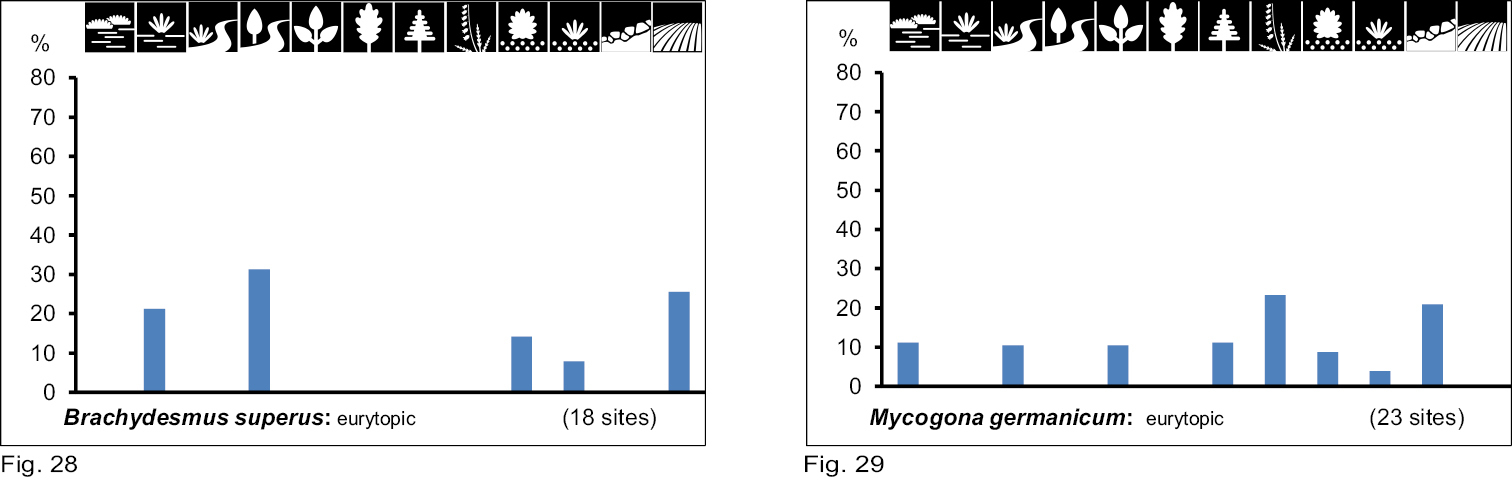
Occurrence of the species of group VII and VIII in the 12 biotope types investigated. Number of positive samples are given in brackets.
Mycogona germanica in group VIII shows no tendency to prefer any biotope type (Table 2, Fig. 29).
Distribution of the diplopod families among inhabitant groupsWith the exception of Glomeridae, millipede families did not show a preference for a particular group (Table 2). In Polydesmidae and Blaniulidae each species investigated belongs to a different group. In Julidae five species occur in very dry and dry biotope types (group VI, e.g. xeric/mesoxeric/ meadows) with single-level vegetation cover, whereas very wet biotope types with multi-level vegetation cover (group II, e.g. floodplains) are inhabited by two species. Only one to three julid species belong to the four other groups (I, III, IV and V; Table 2).
DiscussionAll species of group I (different moisture levels/multi-level vegetation) prefer biotope types without direct insolation due to a multi-level tree and/or shrub layer. Literature data characterise these species as eurytopic woodland species or ubiquists (e.g.
The Main Preferences of the species belonging to group II (very wet/multi-level vegetation) agree with data from the literature (
Craspedosoma rawlinsii Leach, 1815 (Central European). Craspedosoma rawlinsii is one of the few species for which autecological studies exist.
Julus scanicus Lohmander, 1925 (Central European with two distinct south/north western distribution areas and an isolated area in Latvia and Belarus). Especially in its southern distribution area, this species occurs mostly in floodplains and humid deciduous woodlands (
Within group III (wet to humid/multi-level vegetation) we found species which were characterised as eurytopic or woodland species, most abundant in woodland litter and decaying wood (
Tachypodoiulus niger(Leach, 1815) (North West European). Experimental results are that Tachypodoiulus niger is a dry-resistant, xerophilous and cool temperature-preferring species (
Especially remarkable and worthy of discussion are Leptoiulus belgicus and Megaphyllum projectum kochi. Both species demonstrated a special autecological behaviour in the present investigation. They are the only representatives of groups IV and Vrespectively.
Leptoiulus belgicus(Latzel, 1884) (West European with eastern limit of its distribution area in Central Germany). In Saxony-Anhalt this species is most frequent in dry thermophilous oak forests, in shrub associations and in mesoxeric/xeric meadows. The species shows a conspicuously high fidelity to shrub associations, not only in the present investigation area but also in Thuringia (
Megaphyllum projectum kochi (Verhoeff, 1907) (South-East European with north-western limits of its distribution area in Germany). According to
Group VI (very dry to dry/single-level vegetation) contains open land species which are stenotopic (Kryphioiulus occultus, Megaphyllum unilinatum, Choneiulus palmatus and Ophyiulus pilosus) as well as eurytopic (Cylindroiulus caeruleocinctus, Polydesmus inconstans and Ommatoiulus sabulosus). With the exception of Cylindroiulus caeruleocinctus and Ommatoiulus sabulosus all these species will be discussed.
Kryphioiulus occultus (C. L. Koch, 1847) (Eastern European). Over its whole distribution area Kryphioiulus occultus shows a change in habitat preference from south to north. In southern parts (Romania, Hungary) it lives in shrub associations and thermophilous woodlands (Quercetum pubescentis) (
Megaphyllum unilineatum (C. L. Koch, 1838) (East European). This extreme xerothermic species is typical for open sites in its whole distribution area, and also inhabits the south-eastern open woodlands with their dry and warm microclimate (
Ophyiulus pilosus(Newport, 1842) (Northwestern and Central European). Within its northern distribution area (its distribution is disjunct – see
Choneiulus palmatus (Němec, 1895) (European). In Central Europe the species is nearly exclusively synanthropic. German records from near-natural sites are the exception and come exclusively from warm regions (e.g. Rhine-Main-area –
Polydesmus inconstans Latzel, 1884 (Central European). This species shows variable ecological behaviour over its distribution area. At the western boundary (Western Germany, the Netherlands) Polydesmus inconstans is very rare and occurs exclusively at synanthropic localities (
Brachydesmus superus Latzel, 1884 (European). Originally in forests, especially in deciduous woodland (
Mycogona germanica (Verhoeff, 1892) (Central European). This species occurs primarily in the Central European Uplands (Germany, Czech Republic, Poland, Switzerland, Belgium, the Netherlands) with a clear preference for woodland (spruce forests). In Saxony-Anhalt it is limited to the Harz mountain range and its foreland. There the species inhabits bogs, gravel areas, dwarf-shrubs heaths, mesoxeric meadows and other more open habitats in addition to woodlands (
Studies onecological preferences of soil animals can yield different results for one and the same species if they are based on too small a number of replicates. This is due to the fact that many species have the potential to tolerate unfavourable conditions. They can live in “bad” habitats but they do not prefer them (e. g. Craspedosoma). Only investigations based on a large number of observations as in the present study (27, 000 individuals from 175 sites), concentrated on a relatively small investigation area, may yield an approximation to reality. To focus such investigations on a smaller area, at first, seems necessary because the influence of the climatic region on the ecological behaviour of the species should be taken into consideration. Convincing examples for such changes in preferences of chilopod species when considering a larger area of distribution are Cryptops anomalans, Geophilus electricus and Strigamia acuminata (
In small-scale area studies, e.g. in a comparison of different regions within Germany (
I am very grateful to Dr. Peer Schnitter/Halle (Environmental Protection State Office Saxony-Anhalt) for the long-standing cooperative collaboration and the provision of myriapod material. Especially thanks I would like to give to Prof. Wolfram Dunger/Ebersbach and Peter Decker/Görlitz for critical and helpful remarks on the manuscript. Edgar Josephsen/Camano Island took kindly over a first English correction. I also thank him sincerely. I also give many thanks to the editors and referees for additional helpful criticism and English corrections.